
Originalpublikation 2007
Nachstehend der Text in deutscher Übersetzung (Deepl)
Zusammenfassung der Autoren
Es gibt Kontroversen über die Art der Störung in der Gehirnentwicklung, die der Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung (ADHS) zugrunde liegt. Insbesondere ist unklar, ob die Störung auf eine Verzögerung der Gehirnreifung zurückzuführen ist oder ob sie eine vollständige Abweichung von der Vorlage der typischen Entwicklung darstellt. Mithilfe rechnergestützter neuroanatomischer Verfahren haben wir die kortikale Dicke an >40.000 zerebralen Punkten von 824 Magnetresonanztomographien geschätzt, die prospektiv bei 223 Kindern mit ADHS und 223 Kontrollpersonen mit typischer Entwicklung durchgeführt wurden. Mit dieser Stichprobengröße konnten wir den Wachstumsverlauf jedes kortikalen Punktes definieren und eine Phase des Anstiegs in der Kindheit abgrenzen, gefolgt von einer Abnahme der kortikalen Dicke im Jugendalter (ein quadratisches Wachstumsmodell). Aus diesen Verläufen wurde das Alter abgeleitet, in dem die maximale kortikale Dicke erreicht wurde, und als Index für die kortikale Reifung verwendet. Wir stellten fest, dass die Reifung bei Kindern mit und ohne ADHS regional in ähnlicher Weise voranschreitet, wobei die primären sensorischen Areale vor den polymodalen Assoziationsarealen höherer Ordnung die maximale kortikale Dicke erreichen. Allerdings gab es bei ADHS-Kindern eine deutliche Verzögerung beim Erreichen der maximalen Dicke im gesamten Großhirn: Das Medianalter, bei dem 50 % der kortikalen Punkte die maximale Dicke für diese Gruppe erreichten, lag bei 10,5 Jahren (SE 0,01), was deutlich später war als das Medianalter von 7,5 Jahren (SE 0,02) bei sich normal entwickelnden Kontrollpersonen (Log-Rank-Test χ(1)2 = 5.609, P < 1,0 × 10-20). Die Verzögerung war in präfrontalen Regionen, die für die Steuerung kognitiver Prozesse wie Aufmerksamkeit und Bewegungsplanung wichtig sind, am stärksten ausgeprägt. Über eine Verzögerung der regionalen kortikalen Reifung bei ADHS wurde bisher noch nicht neuroanatomisch berichtet.
Die Studie
Die Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung (ADHS) ist die häufigste neurologische Entwicklungsstörung im Kindesalter und betrifft zwischen 3 % und 5 % der Kinder im schulpflichtigen Alter (1). Seit den frühesten Beschreibungen dieser Störung wird darüber diskutiert, ob sie teilweise auf eine verzögerte Hirnreifung zurückzuführen ist oder eine vollständige Abweichung von der Vorlage der typischen Entwicklung darstellt (2). Mehrere Studien kommen zu dem Ergebnis, dass die Gehirnaktivität im Ruhezustand und als Reaktion auf kognitive Tests bei Kindern mit ADHS und ihren etwas jüngeren, aber sich typischerweise entwickelnden Altersgenossen ähnlich ist, was auf eine verzögerte kortikale Entwicklung hindeutet (3–5). Andere berichten jedoch von einer quantitativ unterschiedlichen Neurophysiologie mit einer einzigartigen Architektur des Elektroenzephalogramms und einigen stark anomalen Befunden in funktionellen Bildgebungsstudien, was eher mit ADHS als Abweichung von der typischen Entwicklung übereinstimmt (6–10).
In einer früheren Längsschnittstudie stellten wir bei Kindern mit ADHS und Kontrollpersonen mit typischer Entwicklung parallele Verläufe der Volumenänderung der grauen Lappen fest, aber stärker fokale Veränderungen der kortikalen Reifung, die auf sublobärer Ebene auftreten, würden durch diese lobare Messung nicht erkannt werden (11). Daher wollten wir den Verlauf der kortikalen Entwicklung mithilfe einer Messung der kortikalen Dicke definieren, die eine hervorragende räumliche Auflösung bietet. Die kortikale Dicke wurde als Messgröße gewählt, die sowohl die säulenförmige Architektur des Kortex erfasst als auch empfindlich auf Entwicklungsveränderungen in sich typischerweise entwickelnden und klinischen Populationen reagiert (12–15).
Die meisten der 446 Kinder in der aktuellen Studie wurden wiederholt neuroanatomisch untersucht – 112 (25 %) wurden zweimal, 88 (20 %) dreimal und 30 (7 %) viermal oder öfter gescannt, wobei der durchschnittliche Abstand zwischen den Scans 2,8 Jahre betrug. Solche Längsschnittdaten können mit Querschnittsdaten kombiniert werden, indem eine gemischte Modellregression zur Modellierung von Entwicklungsveränderungen verwendet wird, wobei die Längsschnittdaten besonders aussagekräftig sind. Für Daten zur kortikalen Dicke ist die einfachste Trajektorie, die zur Beschreibung ihrer Veränderung im Laufe der Zeit angepasst werden kann, eine gerade Linie. Komplexere Wachstumsmodelle beinhalten verschiedene Phasen der Zunahme und Abnahme der kortikalen Dicke: Ein quadratisches Modell hat zwei solcher Phasen (in der Regel eine anfängliche Zunahme, die einen Höhepunkt erreicht, bevor sie abnimmt) und ein kubisches Modell hat drei. Abgeleitete Eigenschaften dieser Entwicklungskurven werden häufig als Entwicklungsindizes verwendet, wie z. B. das Alter, in dem Wendepunkte in der Kurve erreicht werden (16, 17). Bei der Betrachtung der kortikalen Veränderung erweist sich das Alter, in dem die maximale kortikale Dicke erreicht wird – der Punkt, an dem die Zunahme der kortikalen Dicke in eine Abnahme übergeht – als besonders nützlicher Index. Beachten Sie, dass die Fähigkeit, ein quadratisches oder kubisches Wachstumsmodell zu erkennen, eine Voraussetzung für die Bestimmung des Alters der maximalen kortikalen Dicke ist; sie kann nicht anhand eines linearen Modells bestimmt werden.
Wir haben daher das Alter, in dem die maximale kortikale Dicke bei Kindern mit und ohne ADHS erreicht wird, verglichen, um festzustellen, ob die Störung durch eine Verzögerung der Reifung der Großhirnrinde gekennzeichnet ist.
Ergebnisse
Die zeitliche Abfolge der kortikalen Reifung, die sich im Alter widerspiegelte, in dem die kortikale Dicke an kortikalen Punkten, für die ein quadratisches Modell geeignet war, ihren Höhepunkt erreichte, war in beiden Gruppen ähnlich [siehe Zusatzinformationen (SI) Filme 1 und 2 und Abb. 1]. Im frontalen Kortex erreichten die superioren, präzentralen und polaren Regionen früh einen Höhepunkt in der kortikalen Dicke, gefolgt von einer zentripetalen Welle, die sich in Richtung des mittleren präfrontalen Kortex bewegte. Im Temporalcortex reiften die hinteren Teile des mittleren und oberen Temporalcortex vor den weiter vorne liegenden Temporalregionen. Im Okzipitalcortex gab es sowohl bei den sich normal entwickelnden als auch bei den ADHS-Probanden frühe Spitzenwerte mit geringen Entwicklungsveränderungen in dem untersuchten Alterszeitraum. Ein direkter Vergleich der kortikalen Veränderungen in den Parietalregionen war schwierig, da sich die Gruppen in den Regionen unterschieden, in denen ein quadratisches Modell angemessen war.
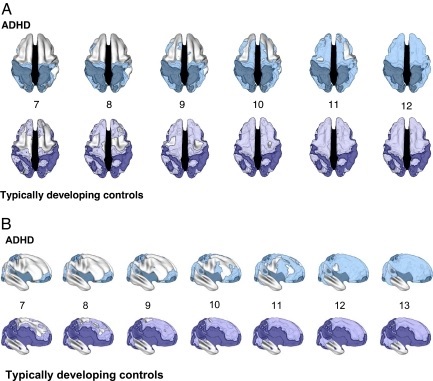
Abb. 1. Das Alter, in dem die maximale kortikale Dicke bei Kindern mit ADHS erreicht wird, im Vergleich zu Kindern mit typischer Entwicklung. (A) Dorsale Ansicht der kortikalen Regionen, in denen die maximale Dicke in jedem Alter (dargestellt im Alter von 7–12 Jahren) bei ADHS (oben) und bei Kontrollpersonen mit typischer Entwicklung (unten) erreicht wurde. Die dunkleren Farben kennzeichnen Regionen, in denen ein quadratisches Modell nicht geeignet war (und daher kein Höchstalter berechnet werden konnte), oder das Höchstalter wurde so geschätzt, dass es außerhalb des abgedeckten Altersbereichs liegt.
Beide Gruppen zeigten eine ähnliche Abfolge der Regionen, die die größte Dicke erreichten, aber die ADHS-Gruppe zeigte eine erhebliche Verzögerung beim Erreichen dieses Entwicklungsmarkers. (B) Rechte Seitenansicht der kortikalen Regionen, in denen die größte Dicke in jedem Alter (dargestellt, Alter 7–13) bei ADHS (oben) und bei sich typischerweise entwickelnden Kontrollpersonen (unten) erreicht wurde. Auch hier ist die Verzögerung der ADHS-Gruppe beim Erreichen der maximalen kortikalen Dicke offensichtlich. Obwohl das allgemeine Entwicklungsmuster ähnlich war, gab es jedoch deutliche diagnostische Unterschiede im zeitlichen Ablauf. Wenn ein Höchstalter bestimmt werden konnte, erreichte die ADHS-Gruppe diesen Meilenstein im Allgemeinen später als die sich typischerweise entwickelnden Kontrollpersonen; siehe Abb. 2. Die Kaplan-Meier-Kurven zeigten, dass das Medianalter, bei dem 50 % der kortikalen Punkte die maximale Dicke erreicht hatten, in der ADHS-Gruppe bei 10,5 Jahren (SE 0,01) lag, was deutlich später war als das Medianalter von 7 ,5 Jahren (SE 0,02) für die sich typischerweise entwickelnden Kontrollpersonen (Log-Rank-Test χ(1)2 = 5.609, P < 1,0 × 10-20); Abb. 3. Die Unterschiede waren am stärksten im mittleren präfrontalen Cortex ausgeprägt, wo die ADHS-Gruppe ihre maximale Dicke ≈5 Jahre nach den sich typisch entwickelnden Kontrollpersonen erreichte, und in geringerem Maße im oberen präfrontalen und medialen präfrontalen Cortex (mit einem Maximum in der ADHS-Gruppe ≈2 Jahre später). Die Kaplan-Meier-Kurven für die präfrontale Region zeigten, dass beide Gruppen zwar eine ähnliche Rate beim Erreichen der kortikalen Dicke aufwiesen, diese jedoch in der ADHS-Gruppe mit einem Medianalter von 10,4 Jahren (SE 0,02) verzögert war, verglichen bei einer Kontrollgruppe mit typischer Entwicklung lag das Medianalter bei 7,5 Jahren (SE 0,02) (Log-Rank-Test χ(1)2 = 9.599, P < 1,0 × 10-20). Nachfolgend war die Verzögerung bilateral im mittleren und oberen temporalen Kortex vorhanden und erstreckte sich bis zum mittleren Hinterhauptskortex, wobei die ADHS-Gruppe ein Höchstalter von 10,6 Jahren (SE = 0,04 ) und bei den sich normal entwickelnden Kontrollpersonen bei 6,8 Jahren (SE = 0,08) lag (Log-Rank-Test χ(1)2 = 303, P < 1,0 × 10-20).
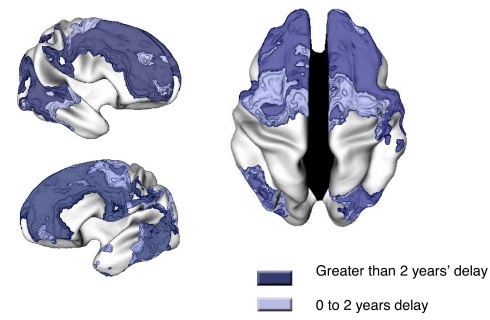 Abb. 2 Regionen, in denen die ADHS-Gruppe eine verzögerte kortikale Reifung aufwies, was durch ein höheres Alter beim Erreichen der maximalen kortikalen Dicke angezeigt wird.
Abb. 2 Regionen, in denen die ADHS-Gruppe eine verzögerte kortikale Reifung aufwies, was durch ein höheres Alter beim Erreichen der maximalen kortikalen Dicke angezeigt wird.
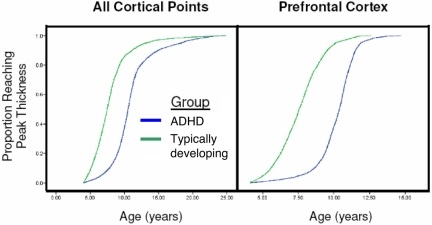 Abb. 3. Kaplan-Meier-Kurven, die den Anteil der kortikalen Punkte veranschaulichen, die in jedem Alter für alle zerebralen kortikalen Punkte (links) und den präfrontalen Kortex (rechts) die maximale Dicke erreicht hatten. Das Medianalter, bei dem 50 % der kortikalen Punkte ihre maximale Dicke erreicht hatten, unterschied sich signifikant zwischen den Gruppen (alle P < 1,0 × 10-20).
Abb. 3. Kaplan-Meier-Kurven, die den Anteil der kortikalen Punkte veranschaulichen, die in jedem Alter für alle zerebralen kortikalen Punkte (links) und den präfrontalen Kortex (rechts) die maximale Dicke erreicht hatten. Das Medianalter, bei dem 50 % der kortikalen Punkte ihre maximale Dicke erreicht hatten, unterschied sich signifikant zwischen den Gruppen (alle P < 1,0 × 10-20).
Die ADHS-Gruppe hatte eine frühere maximale Dicke, vorwiegend im primären motorischen Kortex, mit einem Medianalter, bei dem 50 % der Punkte innerhalb dieser Region bei 7 Jahren (SE = 0,16) lag, verglichen mit 7,4 Jahren (SE 0,12) bei den sich typischerweise entwickelnden Kontrollpersonen (Log-Rank-Test χ(1)2 = 10, P < 0,001);
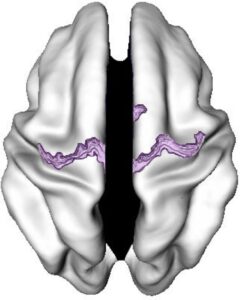 Abb. 4. Regionen, in denen die ADHS-Gruppe eine frühe kortikale Reifung aufwies, was durch ein jüngeres Alter beim Erreichen der maximalen kortikalen Dicke angezeigt wird. Das Ergebnismuster blieb bestehen, wenn der Grad des Bewegungsartefakts in die Regressionsgleichung eingegeben wurde (siehe SI-Abbildungen 5 und 6).
Abb. 4. Regionen, in denen die ADHS-Gruppe eine frühe kortikale Reifung aufwies, was durch ein jüngeres Alter beim Erreichen der maximalen kortikalen Dicke angezeigt wird. Das Ergebnismuster blieb bestehen, wenn der Grad des Bewegungsartefakts in die Regressionsgleichung eingegeben wurde (siehe SI-Abbildungen 5 und 6).
Diskussion
Die kortikale Entwicklung bei Kindern mit ADHS hinkte der Entwicklung bei Kindern mit typischer Entwicklung um mehrere Jahre hinterher. Die geordnete Abfolge der regionalen Entwicklung, bei der die primären sensorischen und motorischen Bereiche ihre maximale kortikale Dicke vor den höherwertigen Assoziationsbereichen erreichen, war jedoch in beiden Gruppen ähnlich, was darauf hindeutet, dass ADHS eher durch eine Verzögerung als durch eine Abweichung in der kortikalen Reifung gekennzeichnet ist. Dies steht im Gegensatz zu anderen neurologischen Entwicklungsstörungen wie Autismus, bei denen es zu einer dramatischen Verschiebung der Gehirnwachstumskurven entlang der Altersachse nach rechts zu kommen scheint, was dazu führt, dass das maximale Gehirnvolumen in einem viel früheren Alter erreicht wird – das Gegenteil des Musters, das wir bei ADHS beobachten (18, 19).
Die Verzögerung der kortikalen Reifung bei ADHS war am ausgeprägtesten im lateralen präfrontalen Kortex, der Region mit den konsistentesten Berichten über strukturelle Anomalien bei dieser Störung (11, 20), insbesondere in den superioren und dorsolateralen präfrontalen Regionen (21–23). Der präfrontale Kortex unterstützt eine Vielzahl kognitiver Funktionen, wie die Fähigkeit, unangemessene Reaktionen und Gedanken zu unterdrücken (24, 25), die exekutive „Kontrolle“ der Aufmerksamkeit (26), die Bewertung von Belohnungskontingenzen (27, 28), die motorische Kontrolle höherer Ordnung (5) und das Arbeitsgedächtnis (29). Defizite in diesen kognitiven Funktionen wurden alle mit der Pathogenese von ADHS in Verbindung gebracht (30), und eine präfrontale kortikale Hypoaktivierung bei Kindern mit ADHS während der Ausführung vieler dieser Aufgaben ist ein relativ konsistenter Befund (10).
Eine Verzögerung wurde auch im temporalen Kortex festgestellt, am deutlichsten in den hinteren Abschnitten des mittleren/oberen temporalen Gyrus bilateral, relativ umschrieben auf der linken Seite und mit einer weiter hinten liegenden Ausdehnung auf der rechten Seite. Strukturelle Veränderungen in den Temporallappen sind ein häufiger Befund in Studien zu ADHS, von der Ebene des gesamten Lappens (11) bis hin zu stärker fokalen Anomalien der Dichte der grauen Substanz und der kortikalen Dicke (31, 32), und können metabolische (9, 33), funktionelle (10, 34–36) und elektrophysiologische Korrelate (37, 38) aufweisen. Ein gemeinsames Merkmal der Frontal- und Temporalregionen mit der größten Reifeverzögerung ist die Beteiligung des heteromodalen Kortex (39). Dabei handelt es sich um miteinander verbundene kortikale Regionen, die Informationen aus sensorischen Bereichen niedrigerer Ordnung integrieren und Wahrnehmungen höherer Ordnung erzeugen, die die Steuerung von Aufmerksamkeit und Handlung steuern. Strukturelle Anomalien dieses Systems wurden mit der Pathogenese von ADHS in Verbindung gebracht (31).
Im Gegensatz dazu war der primäre motorische Kortex der einzige kortikale Bereich, in dem die ADHS-Gruppe eine etwas frühere Reifung aufwies. Es ist möglich, dass die Kombination aus früher Reifung des primären motorischen Cortex und später Reifung der übergeordneten motorischen Kontrollregionen die exzessive und schlecht kontrollierte motorische Aktivität widerspiegelt oder sogar antreibt, die für das Syndrom kennzeichnend ist.
Das Erreichen der maximalen kortikalen Dicke in einem jüngeren Alter bedeutet auch, dass die sich typischerweise entwickelnden Kinder früher in die Phase der kortikalen Ausdünnung eintreten, die die Adoleszenz dominiert (40, 41). Aufgrund der begrenzten Altersspanne konnten wir nicht das Alter definieren, in dem die jugendliche Phase der kortikalen Ausdünnung abflacht und in stabile kortikale Dimensionen bei Erwachsenen übergeht. Wir gehen davon aus, dass das Alter, in dem diese im Wesentlichen statische Erwachsenenphase erreicht wird, bei den Probanden mit ADHS ebenfalls später liegt.
Unseres Wissens nach wurden bisher keine neuroanatomischen Belege für die Theorie einer Verzögerung der kortikalen Reifung bei ADHS berichtet. Die Verwendung einer kortikalen Messung, die eine exquisite räumlich-zeitliche Auflösung bietet, ermöglicht es uns, eine erhebliche Variabilität im zeitlichen Ablauf der kortikalen Reifung innerhalb jedes Lappens nachzuweisen, die durch unsere früheren volumetrischen Lappenanalysen nicht erkennbar war (11). Darüber hinaus können wir die größte Reifeverzögerung in präfrontalen kortikalen Regionen lokalisieren, die an der Pathogenese von ADHS beteiligt sind.
In anderen Arbeiten an einer Teilstichprobe von Probanden mit klinischen Ergebnisdaten aus dieser Kohorte konnten wir nur lineare Veränderungsmuster in der kortikalen Dicke feststellen (und konnten daher das Alter der maximalen kortikalen Dicke nicht definieren) und fanden im Allgemeinen parallele Verläufe mit Ausnahme einer Region im rechten parietalen Kortex (12). Durch die Einbeziehung zusätzlicher Probanden können wir Effekte höherer Ordnung des Alters erkennen und somit diagnostische regionale Unterschiede im Alter des Erreichens der maximalen kortikalen Dicke (12) herausarbeiten. Da uns in der aktuellen Studie klinische Ergebnisdaten für die Mehrheit der ADHS-Probanden fehlten, konnten wir nicht untersuchen, ob ein gutes oder schlechtes klinisches Ergebnis mit Unterschieden im Zeitpunkt wichtiger Entwicklungsmarker wie dem Alter der maximalen kortikalen Dicke zusammenhängt.
Um auf die zentrale Erkenntnis zurückzukommen: Das allgemein höhere Alter, in dem die maximale kortikale Dicke bei ADHS erreicht wird, stellt vermutlich eine zeitliche Verschiebung im Gleichgewicht zwischen den zellulären Prozessen dar, die zu einer anfänglichen Zunahme und späteren Abnahme der kortikalen Dicke führen. Die genaue Art dieser Prozesse bei sich normal entwickelnden Kindern muss noch bestimmt werden. Aus Tierversuchen lässt sich ableiten, dass die Zunahme der kortikalen Dicke durch Mechanismen wie das Wachstum der dendritischen Dornen und die Ausarbeitung von Stützglia und Gefäßen verursacht werden könnte (42, 43). Die kortikale Ausdünnung im Jugendalter könnte auf eine intrakortikale Myelinisierung und die nutzungsabhängige selektive Beseitigung von Synapsen zurückzuführen sein, die zur Bildung und Gestaltung neuronaler Schaltkreise beitragen könnten, einschließlich solcher, die kognitive Fähigkeiten unterstützen (44–46). Was ADHS betrifft, so basieren Tiermodelle meist auf Störungen der monoaminergen Neurotransmission, die als Reaktion auf frühe Schädigungen (z. B. induzierte vorübergehende Hyperthyreose oder neonatale 6-OHDA-Läsionen) oder Anomalien von Neurotransmittern (wie die 160-bp-Insertion in Exon 3 des Dopamintransportgens bei der spontan hypertensiven Ratte) auftreten (47, 48). Wie solche Veränderungen die Dynamik der kortikalen Entwicklung beeinflussen könnten, ist noch unklar, wäre aber ein wichtiges Gebiet für zukünftige Forschung.
Welche ätiologischen Faktoren könnten dieser Verzögerung zugrunde liegen? Trophische Effekte der Behandlung mit Psychostimulanzien in der ADHS-Gruppe sind möglich, aber unwahrscheinlich, da wir in unseren früheren Berichten keine Auswirkungen von Psychostimulanzien auf das Volumen der grauen Substanz festgestellt haben (11). Da es sich bei unseren Studien jedoch um Beobachtungsstudien handelt, sind alle Schlussfolgerungen zu Stimulanzien vorläufig. Unsere Gesamtergebnisse können nicht auf Gruppenunterschiede in Bezug auf Intelligenz und Geschlecht zurückgeführt werden, die zwar die kortikale Entwicklung beeinflussen (14, 41, 49, 50), aber in unserem Design streng kontrolliert wurden. Genetische Faktoren werden sicherlich eine Rolle spielen, mit einer Störung in der Entwicklungssequenz der Aktivierung und Deaktivierung von Genen, die die kortikale Architektur formen. In diesem Zusammenhang erweisen sich Neurotrophine, die für die Proliferation, Differenzierung und das Überleben neuronaler und nicht-neuronaler Zellen unerlässlich sind, als vielversprechende Kandidaten, und tatsächlich wurden Polymorphismen innerhalb der Gene für den hirneigenen neurotrophen Faktor und den Nervenwachstumsfaktor 3 bereits versuchsweise mit ADHS in Verbindung gebracht (51, 52).
Verlaufsstudien zur Gehirnentwicklung, die auf neuroanatomischen Längs- und Querschnittsdatensätzen basieren, liefern wertvolle Erkenntnisse über ADHS. Sie liefern nicht nur Informationen für wichtige Debatten, die seit den frühesten Beschreibungen der Störung (2) geführt werden, sondern können auch die zukünftige Suche nach Faktoren leiten, die die kortikale Entwicklung verzögern, anstatt sie zu beeinträchtigen.
Methoden
Probanden
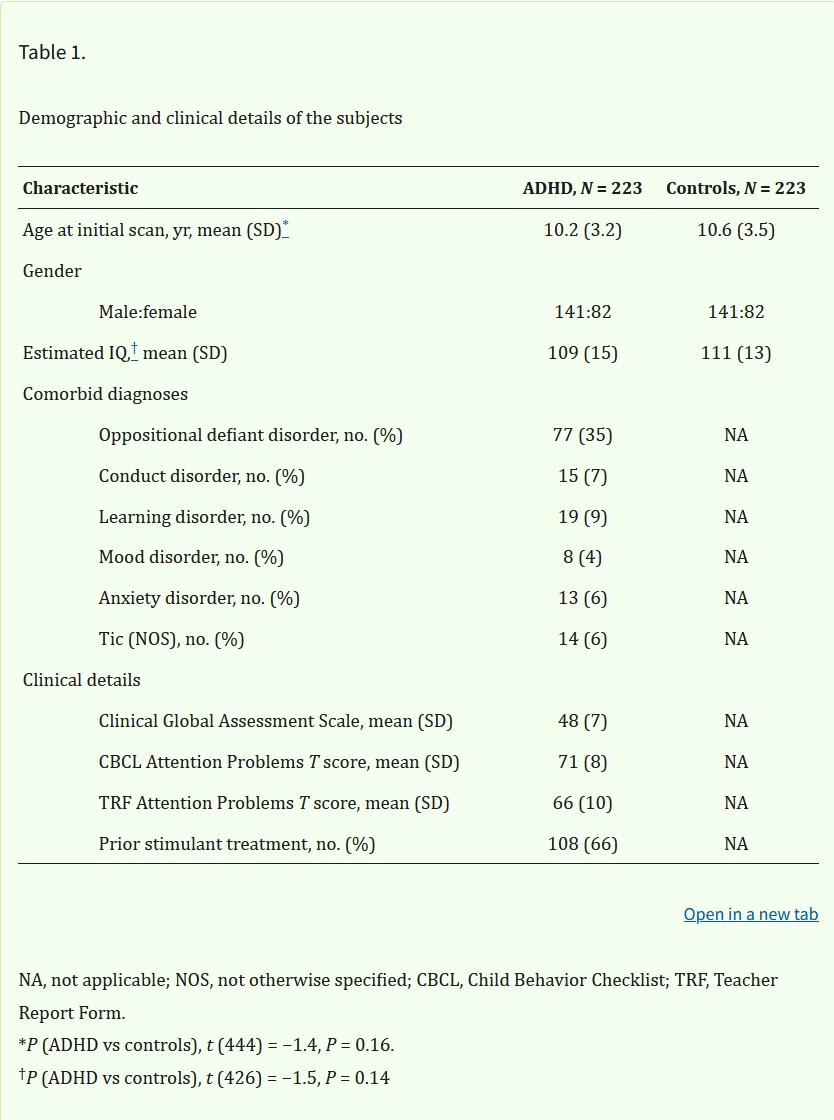
Die klinische Gruppe umfasste 223 Kinder und Jugendliche mit ADHS gemäß der Definition des Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition (DSM-IV). Die Diagnose basierte auf dem Parent Diagnostic Interview for Children and Adolescents (53), den Conner’s Teacher Rating Scales (54) und dem Teacher Report Form; siehe Tabelle 1. Ausschlusskriterien waren ein IQ unter 80 und Anzeichen für medizinische oder neurologische Störungen. 205 (92 %) hatten zu Studienbeginn eine kombinierte Form von ADHS, 13 (6 %) eine unaufmerksame Form und 5 (2 %) eine hyperaktive/impulsive Form. 154 nicht verwandte Einzelkinder und 25 betroffene Geschwisterpaare (mit 53 Personen) sowie 16 Zwillingsgeburten (nur ein Kind pro Zwillingspaar) wurden einbezogen. Es wurden Kontrollpersonen mit typischer Entwicklung rekrutiert, und jede Person füllte die Childhood Behavior Checklist als Screening-Instrument aus und unterzog sich dann einem strukturierten diagnostischen Interview durch einen Kinderpsychiater, um psychiatrische oder neurologische Diagnosen auszuschließen (55). Die Teilnehmer mit typischer Entwicklung in dieser Studie wurden in Bezug auf Geschlecht, Alter und Intelligenz, gemessen anhand einer altersgerechten Version der Wechsler-Intelligenztests, mit der ADHS-Gruppe abgeglichen. Es gab 169 Einzelkinder, 17 Gruppen nicht betroffener Geschwister (mit 38 Personen) und 16 Zwillingsgeburten (ein Kind pro Zwillingspaar). Die Ethikkommission der National Institutes of Health genehmigte das Forschungsprotokoll, und von den Eltern bzw. Kindern wurde eine schriftliche Einverständniserklärung und Zustimmung zur Teilnahme an der Studie eingeholt.
Tabelle 1. Demografische und klinische Details der Probanden
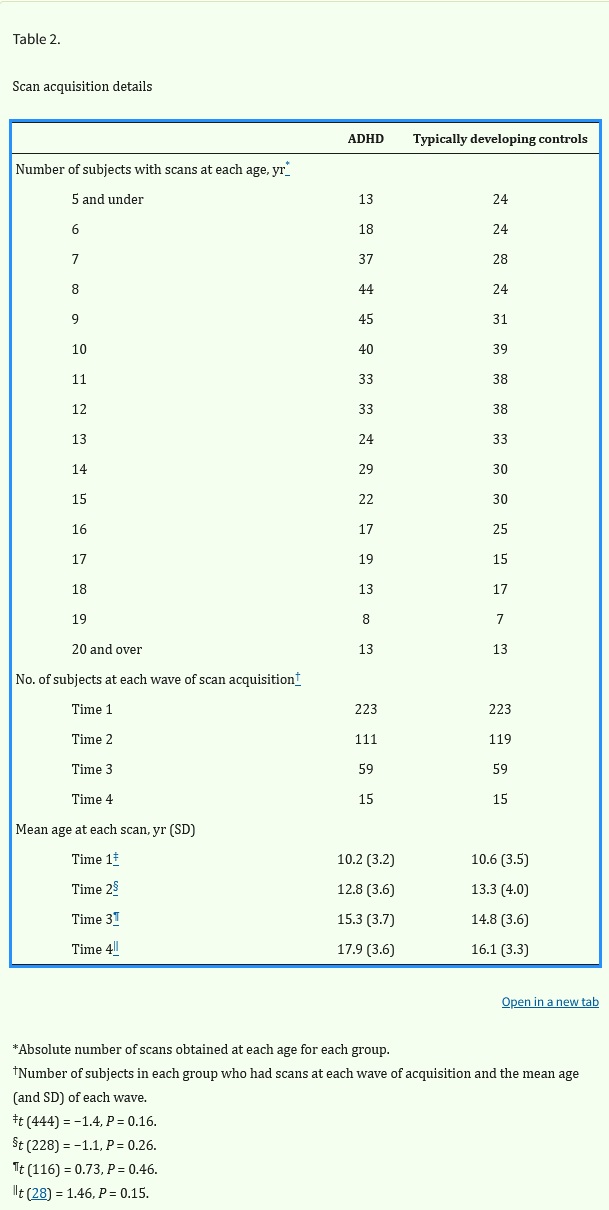
Die Gesamtzahl der Probanden, die in jedem Alter gescannt wurden, ist in Tabelle 2 angegeben. Dort ist auch die Anzahl der Probanden aufgeführt, die wiederholt gescannt wurden, sowie das Durchschnittsalter bei jeder Scan-Welle. Das durchschnittliche Intervall zwischen den Scans betrug 2,9 Jahre (SD 1,5) für die ADHS-Gruppe und 2,8 Jahre (SD 1,4) für die Kontrollgruppe mit typischer Entwicklung [t (317) = 1,3, P = 0,2].
Tabelle 2. Details zur Scanerfassung
Neuroimaging
Alle Kinder wurden während der gesamten Studie mit einem 1,5-T-General-Electric-Signa-Scanner einer neuroanatomischen Magnetresonanztomographie unterzogen. Die Bildgebungsparameter waren eine Echozeit von 5 ms, eine Wiederholungszeit von 24 ms, ein Flipwinkel von 45°, eine Erfassungsmatrix von 256 × 192, eine Anregungszahl von 1 und ein Sichtfeld von 24 cm. Die Kopfpositionierung wurde wie beschrieben standardisiert (56). Für die gesamte Studie wurde derselbe 1,5-T-Signa-Scanner von General Electric verwendet. Die nativen MRT-Scans wurden mithilfe einer linearen Transformation in einem standardisierten stereotaktischen Raum registriert und um Artefakte aufgrund von Ungleichförmigkeit korrigiert (57). Die registrierten und korrigierten Volumina wurden mithilfe eines fortschrittlichen neuronalen Netzklassifikators (58) in weiße und graue Substanz, Liquor und Hintergrund unterteilt. Die innere und äußere kortikale Oberfläche wurden dann mithilfe verformbarer Modelle extrahiert und nichtlinear an einer Standard-Schablonenoberfläche ausgerichtet (59). Die kortikale Dicke wurde dann im nativen Raum in Millimetern gemessen, indem der verknüpfte Abstand zwischen den Oberflächen der pialen weißen und grauen Substanz an 40.960 Scheitelpunkten in der gesamten Großhirnrinde verwendet wurde. Bei der Schätzung der kortikalen Dicke wählten wir auf der Grundlage einer Populationssimulationsstudie einen Unschärfekern mit einer Bandbreite von 30 mm, der zeigte, dass diese Bandbreite die statistische Aussagekraft maximiert und gleichzeitig falsch positive Ergebnisse minimiert (60). Dieser Kern bewahrt die Fähigkeit zur anatomischen Lokalisierung, da eine Unschärfe von 30 mm entlang der Oberfläche unter Verwendung eines Diffusionsglättungsoperators die kortikalen topologischen Merkmale bewahrt und deutlich weniger Kortex darstellt als der äquivalente volumetrische Gaußsche Unschärfekern (60). Alle Scans wurden nach dem Grad des Bewegungsartefakts (kein, leichtes, mittleres oder schweres Bewegungsartefakt) bewertet, wie in SI Text und in Ref. 61 beschrieben. Scans mit mittlerem oder schwerem Bewegungsartefakt wurden von weiteren Analysen ausgeschlossen; Scans mit leichtem Bewegungsartefakt wurden einbezogen.
Statistische Analysen
Zunächst bestimmten wir die Entwicklungsverläufe mithilfe einer gemischten Modellregressionsanalyse, die die Einbeziehung mehrerer Messungen pro Person, fehlender Daten und unregelmäßiger Intervalle zwischen den Messungen ermöglicht und so die statistische Aussagekraft erhöht (62). Ein Zufallseffekt für jede Person wurde in einen Zufallseffekt für jede Familie eingebettet, wodurch sowohl die Abhängigkeit innerhalb der Person als auch innerhalb der Familie berücksichtigt wurde. Unsere Klassifizierung der Entwicklungsverläufe basierte auf einem Auswahlverfahren für ein Step-down-Modell: An jedem kortikalen Punkt modellierten wir die kortikale Dicke mithilfe eines Polynom-Regressionsmodells mit gemischten Effekten und testeten auf kubische, quadratische und lineare Alterseffekte. Wenn der kubische Alterseffekt bei P < 0,05 nicht signifikant war, wurde er entfernt, und wir gingen zum quadratischen Modell über und so weiter. Auf diese Weise konnten wir die Entwicklung jedes kortikalen Punktes so klassifizieren, dass sie am besten durch eine kubische, quadratische oder lineare Funktion des Alters erklärt werden konnte. Ein quadratisches Modell erwies sich für einen Großteil des Kortex als geeignet, wobei die k-te kortikale Dicke des i-ten Individuums in der j-ten Familie wie folgt modelliert wurde: Dickeijk = Achsenabschnitt + dij + β1(Alter – Durchschnittsalter) + β2*(Alter – Durchschnittsalter)**2) + eijk, wobei dij verschachtelte Zufallseffekte sind, die die Abhängigkeit innerhalb der Person und innerhalb der Familie modellieren, die Achsenabschnitts- und β-Terme feste Effekte sind und eijk den Restfehler darstellt. Insbesondere war sowohl für die ADHS- als auch für die Kontrollgruppe mit typischer Entwicklung ein quadratisches Modell für den größten Teil des lateralen präfrontalen und medialen präfrontalen Kortex, des oberen und mittleren temporalen Kortex, des oberen und mittleren okzipitalen Kortex sowie der angularen und supramarginalen Gyri geeignet. Die ADHS-Gruppe zeigte im Gegensatz zu den Kontrollpersonen mit typischer Entwicklung, für die ein quadratisches Modell galt, eine lineare Anpassung in den oberen Parietallappen und den postzentralen Gyri. Die Analysen wurden wiederholt, wobei der Grad des Bewegungsartefakts in die Regressionsgleichung eingegeben wurde. Als Nächstes wurde das Alter, in dem die maximale kortikale Dicke für jede Gruppe erreicht wurde, in diesen Regionen aus den Ableitungen erster Ordnung der angepassten Kurven berechnet und durch dynamische Zeitraffersequenzen („Filme“) veranschaulicht. Es wurden Kaplan-Meier-Kurven erstellt, die den Anteil der kortikalen Punkte zeigen, die im gesamten abgedeckten Altersbereich die maximale kortikale Dicke erreicht hatten. Die Signifikanz des Gruppenunterschieds im Medianalter, bei dem die Hälfte der kortikalen Punkte ihre maximale Dicke erreicht hatte, wurde mithilfe des Log-Rank-Tests (Mantel-Cox-Test) berechnet.
Gehirnkarten zeigen die Regionen, in denen die ADHS-Gruppe entweder in einem früheren oder späteren Alter die größte Dicke aufwies.
In der Originalpublikation folgen hier die Referenzen.