
https://onlinelibrary.wiley.com/doi/full/10.1002/hbm.23401?utm_source=chatgpt.com
Human Brain Mapping, Band 38, Ausgabe 2 pp. 631-643, Forschungsartikel, Offener Zugang
Isabella A. Breukelaar, Cassandra Antees, Stuart M. Grieve, Sheryl L. Foster, Lavier Gomes, Leanne M. Williams, Mayuresh S. Korgaonkar Erstmals veröffentlicht: 13. September 2016 https://doi.org/10.1002/hbm.23401
Vollständiger Titel: Die Anatomie des kognitiven Kontrollnetzes korreliert mit dem neurokognitiven Verhalten: Eine Längsschnittstudie (Cognitive control network anatomy correlates with neurocognitive behavior: A longitudinal study)
Zusammenfassung
Kognitive Kontrolle ist der Prozess des Einsatzes exekutiver Funktionen wie Aufmerksamkeit, Planung oder Arbeitsgedächtnis zur Steuerung geeigneter Verhaltensweisen, um ein bestimmtes Ziel zu erreichen. Studien zur funktionellen Magnetresonanztomographie deuten auf ein übergeordnetes kognitives Kontrollnetzwerk hin, das die dorsalen Regionen des lateralen präfrontalen Kortex (DLPFC), des anterioren cingulären Kortex (dACC) und des parietalen Kortex (DPC) umfasst. Wie sich die Struktur der grauen Substanz in diesem Netzwerk während der Neuroentwicklung verändert und wie sich diese Veränderungen auf die kognitive Kontrolle auswirken, ist noch nicht vollständig geklärt. Hier untersuchen wir die Veränderungen des Volumens der grauen Substanz in den wichtigsten Knotenpunkten des kognitiven Kontrollnetzwerks anhand struktureller MRT-Scans von 176 Teilnehmern im Alter von 8 bis 38 Jahren. Einhundertelf von ihnen schlossen auch eine Längsschnittuntersuchung nach zwei Jahren ab. Wir vergleichen diese Ergebnisse mit der Leistung in einer kognitiven Batterie, die ebenfalls zu diesen beiden Zeitpunkten gemessen wurde. Wir fanden heraus, dass Volumenverringerungen im kognitiven Kontrollnetzwerk mit verbesserten Leistungen bei exekutiven Funktionen (im linken DLPFC und bilateralen DPC), bei der Informationsverarbeitung (im bilateralen dACC und rechten DPC) und bei Aufgaben zur Emotionsidentifikation (linker DLPFC) einhergingen. Diese Ergebnisse waren auch nach Kontrolle des Alters signifikant. Außerdem waren die Veränderungen der grauen Substanz im gesamten Netzwerk koordiniert. Diese Ergebnisse deuten darauf hin, dass altersunabhängiges synaptisches Pruning im kognitiven Kontrollnetzwerk eine Rolle bei der Verbesserung der Leistung in kognitiven Bereichen spielen könnte. Diese Studie gibt Aufschluss über die direkten Auswirkungen struktureller Veränderungen auf das Verhalten innerhalb dieses Netzwerks während der Neuroentwicklung und liefert eine normative Evidenzbasis für ein besseres Verständnis der Entwicklung kognitiver Dysfunktionen bei Hirnstörungen. Hum Brain Mapp 38:631-643, 2017. © 2016 The Authors Human Brain Mapping Published by Wiley Periodicals, Inc.
Abkürzungen
CCN Kognitives Kontrollnetzwerk
dACC Anteriorer cingulärer Kortex
DLPFC Dorsolateraler präfrontaler Kortex
DPC Dorsal/Posteriorer parietaler Kortex
EINFÜHRUNG
Die kognitive Kontrolle ist ein Schlüsselaspekt des normalen Funktionierens, da sie an der Regulierung emotionaler und sozialer Signale und Reaktionen sowie an formalen Intelligenzmaßen wie Gedächtnis und Aufmerksamkeit beteiligt ist. Beeinträchtigungen der kognitiven Kontrolle haben schwerwiegende Auswirkungen auf die Fähigkeit eines Individuums, sich sowohl in seiner inneren als auch in seiner äußeren Welt zurechtzufinden, was durch das konsequente Vorhandensein einer kognitiven Kontrollstörung bei psychiatrischen Erkrankungen belegt wird [Millan et al., 2012; Williams, 2016]. Jüngste Ergebnisse der funktionellen Magnetresonanztomographie (fMRI) legen nahe, dass die kognitive Kontrolle durch die gleichzeitige Aktivierung von Regionen im frontalen und parietalen Kortex aufrechterhalten wird, nämlich dem dorsalen anterioren cingulären Kortex (dACC), dem dorsolateralen präfrontalen Kortex (DLPFC) und dem dorsalen/posterioren parietalen Kortex (DPC). Diese Areale bilden ein Netzwerk, das mehrere Bereiche der kognitiven Kontrolle kontrolliert und koordiniert, darunter Aufmerksamkeit, Arbeitsgedächtnis, Hemmung, Planung und andere exekutive Funktionen als Reaktion auf Aufgabenanforderungen [Niendam et al., 2012]. Ein Verständnis des kognitiven Kontrollnetzwerks (CCN) auf Schaltkreisebene hat ein neues Paradigma dafür geschaffen, wie eine optimale kognitive Kontrollfunktion im gesunden Gehirn erreicht wird und warum kognitive Kontrollstörungen bei psychiatrischen Störungen auftreten [Williams, 2016]. Wenn wir verstehen, wie sich die neuronale Struktur dieses Netzwerks in der späten Kindheit bis zum frühen Erwachsenenalter verändert, einer wichtigen Entwicklungsphase für das Risiko verschiedener psychiatrischer Störungen, und wie diese Veränderungen mit der kognitiven Leistung zusammenhängen, wird dies unser Verständnis des neurologischen Entwicklungsprozesses der kognitiven Kontrolle fördern.
Das Muster der Veränderung des Volumens der grauen Substanz im sich entwickelnden Gehirn wird seit langem als Indikator für die strukturelle Reifung verwendet, aber die Beziehung zwischen dieser und der Verhaltensentwicklung ist nach wie vor schlecht verstanden [Casey et al., 2005]. Man geht davon aus, dass die Veränderung des Volumens der grauen Substanz eine Kombination aus Myelinisierung und synaptischem Pruning während der Neuroentwicklung widerspiegelt und einen Phänotyp für das Verständnis der verschiedenen Rollen vorbestimmter genetischer Hinweise und erfahrungsabhängiger Mechanismen bei der Entwicklung von Funktionen höherer Ordnung liefert. In dieser Studie wollen wir anhand eines Längsschnittdesigns untersuchen, wie der normale Entwicklungsverlauf des Volumens der grauen Substanz in den Schlüsselbereichen des CCN mit dem kognitiven Kontrollverhalten zusammenhängt.
Das Muster der Veränderung der kortikalen grauen Substanz während der Kindheit und Jugend wurde in einer Reihe von Querschnittsstudien [Pfefferbaum et al., 1994; Thompson et al., 2005; Toga et al., 2006] und einigen Längsschnittstudien mit struktureller MRT [Giedd et al., 1999; Gogtay et al., 2004; Lenroot et al., 2007; Shaw et al., 2008; Sowell et al., 2003; Sowell et al., 2004] untersucht und repliziert. Das Volumen der grauen Substanz scheint bis zur Pubertät zuzunehmen, danach beginnt es zu sinken. Dieses Muster ist jedoch im gesamten Gehirn gestaffelt, wobei die sensomotorischen Bereiche zuerst reifen, gefolgt von den Bereichen, die für höhere kognitive Funktionen verantwortlich sind, wie die Frontallappen und die dorsalen Parietallappen [Giedd et al., 1999; Gogtay et al., 2004]. Es wurde festgestellt, dass die Verhaltensmeilensteine einem ähnlichen Muster folgen, wobei die sensomotorischen Fähigkeiten in der späten Kindheit heranreifen und die kognitiven Funktionen höherer Ordnung wie Sprache und Exekutivfunktionen weiterhin durch die Adoleszenz bis ins frühe Erwachsenenalter reifen [Anderson, 2002; Best und Miller, 2010; Clark et al., 2006; Schaie und Strother, 1968; Welsh et al., 1991].
Allerdings ist es nur wenigen Studien gelungen, diese Ereignisse miteinander zu verknüpfen. Die meisten Studien zu Entwicklungsprozessen sind ausschließlich neuroanatomisch oder verhaltensorientiert. Darüber hinaus geht die Mehrzahl der Studien zur Neuroentwicklung von Annahmen aus, die auf Querschnittsdaten beruhen, was ein unvollständiges und potenziell irreführendes Bild der Entwicklungsveränderungen vermittelt [Kraemer et al., 2000]. Nur wenige Längsschnittstudien haben die kortikale Entwicklung mit kognitiven Veränderungen in Verbindung gebracht. Sowell et al. wiesen nach, dass eine stärkere kortikale Ausdünnung in den linken dorsalen frontalen und parietalen Regionen bei Kindern mit großen Verbesserungen bei einer Messung der verbalen Intelligenz verbunden war. Diese Studie war jedoch durch eine kleine Stichprobengröße (n = 45) und eine enge Altersspanne (5-11 Jahre) begrenzt [Sowell et al., 2004]. Eine zweite Studie [Shaw et al., 2006], die eine wesentlich größere Stichprobengröße (n = 307) und eine größere Altersspanne (7-27 Jahre) verwendete, fand heraus, dass die Veränderungsrate der kortikalen Dicke vor allem in den frontalen Regionen mit der Messung der allgemeinen Intelligenz korrelierte, ein Ergebnis, das inzwischen wiederholt wurde [Burgaleta et al., 2014]. Während diese Studien nützliche Verbindungen zwischen Intelligenz und Veränderungen in der kortikalen Struktur herstellen, hat nur eine andere Gruppe Längsschnittveränderungen im Gehirn mit einem bestimmten kognitiven Bereich in Verbindung gebracht. Tamnes et al. fanden heraus, dass verbesserte Leistungen im Arbeitsgedächtnis mit longitudinalen Abnahmen in bilateralen präfrontalen und posterioren parietalen Regionen zusammenhingen [Tamnes et al., 2013]. Wie die Volumina der grauen Substanz mit der Leistung in anderen kognitiven Bereichen wie Aufmerksamkeit, Exekutivfunktion, Effizienz der Informationsverarbeitung und kognitiven Aspekten der Emotionsverarbeitung zusammenhängen, ist noch unklar. Diese Studien befassen sich zwar mit der allgemeinen Veränderung der grauen Substanz mit dem Alter im gesamten Gehirn und haben verschiedene Teile des CCN einbezogen, aber ob diese Veränderung innerhalb des CCN entwicklungsmäßig koordiniert ist, muss noch ermittelt werden. Es gibt Hinweise darauf, dass es koordinierte Muster anatomischer Veränderungen gibt, die mit dem Grad der funktionellen Konnektivität zwischen verschiedenen Bereichen des Gehirns korrelieren [Raznahan et al., 2011], was wir in dieser Studie innerhalb des CCN weiter untersuchen.
Diese Studie zielt darauf ab zu verstehen, wie Veränderungen des Volumens der grauen Substanz in den Schlüsselknoten des CCN miteinander und mit Leistungsveränderungen in sieben wichtigen kognitiven Bereichen während einer entscheidenden Reifungsphase von der späten Kindheit bis ins Jugend- und Erwachsenenalter zusammenhängen. Wir verwenden strukturelle MRT- und kognitive Daten von 176 Teilnehmern im Alter von 8-38 Jahren, von denen 111 auch eine zweijährige longitudinale Nachuntersuchung absolvierten. Wir haben Querschnittsdaten verwendet, um zunächst die Beziehung zwischen dem Alter und den Verhaltensmaßen der kognitiven Kontrolle und der Struktur der grauen Substanz des CCN zu ermitteln und um festzustellen, ob diese im Längsschnitt über die Zeit erhalten bleiben. Außerdem testen wir die Querschnittsbeziehungen zwischen kognitivem Verhalten und der Struktur der grauen Substanz und untersuchen, wie die Veränderung der Gehirnstruktur innerhalb des CCN mit den Veränderungen des kognitiven Verhaltens im Längsschnitt zusammenhängt. Wir stellten die Hypothese auf, dass das Alter eine signifikante Beziehung sowohl zum Volumen der grauen Substanz als auch zur kognitiven Leistung hat. Auf der Grundlage früherer Studien, die sich speziell mit Kognition und Neuroanatomie befassten und/oder ein Querschnittsmodell verwendeten, sagten wir voraus, dass eine Abnahme der grauen Substanz im CCN während der Neuroentwicklung eine Zunahme der kognitiven Fähigkeiten vorhersagt. Außerdem gingen wir davon aus, dass der Verlauf der Veränderung des Volumens der grauen Substanz im gesamten CCN koordiniert sein würde, da es eine zusammenhängende funktionelle Rolle bei der kognitiven Kontrolle spielt.
MATERIALIEN UND METHODEN
Teilnehmer und Studiendesign
Einhundertneunundsiebzig gesunde Teilnehmer im Alter zwischen 8 und 38 Jahren wurden rekrutiert, um im Westmead Hospital (Sydney, Australien) eine strukturelle MRT-Untersuchung sowie eine gut etablierte kognitive Batterie [Mathersul et al., 2009; Williams et al., 2009] am Westmead Institute for Medical Research (Sydney, Australien) durchzuführen. Einhundertfünfzehn dieser Teilnehmer kehrten zu einer Nachuntersuchung nach zwei Jahren zurück, bei der sie das gleiche Protokoll ausfüllten (siehe Abb. 1). Drei Teilnehmer wurden ausgeschlossen, weil sie zum Zeitpunkt der Nachuntersuchung psychisch erkrankt waren und/oder bei ihnen zufällig eine strukturelle Anomalie im Gehirn festgestellt wurde. Ein Teilnehmer wurde wegen des Auftretens von bewegungsbezogenen Artefakten ausgeschlossen.
Abbildung 1 (hier in PowerPoint)
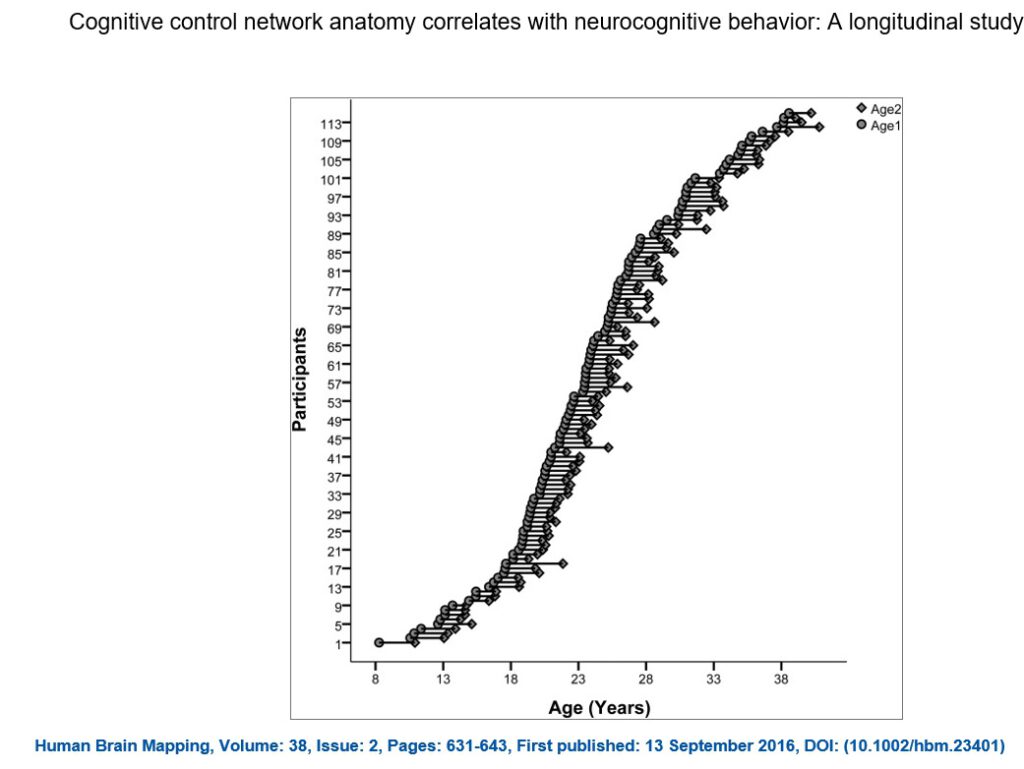
Altersverteilung der Längsschnittdaten (Jahre). Die Teilnehmer im Alter von 8 bis 38 Jahren (n = 111, 53 Frauen) unterzogen sich einer 3T-MRT-Untersuchung und absolvierten eine kognitive Testbatterie zu zwei Zeitpunkten, die im Durchschnitt 1,94 Jahre auseinander lagen. Das Alter bei jedem Scan ist durch einen Kreis gekennzeichnet. Der erste Scan ist der am weitesten links liegende Scan, während der wiederholte Scan direkt rechts liegt und durch eine horizontale Linie verbunden ist.
Die Teilnehmer mussten bei beiden Untersuchungen die Zulassungskriterien erfüllen und durften nach eigenen Angaben keine schwerwiegenden Erkrankungen, keine frühere oder aktuelle Diagnose einer schweren Depression und/oder Suizidalität, einer bipolaren Störung, einer Psychose, einer Schizophrenie, einer schizoaffektiven Störung, einer primären Essstörung, einer posttraumatischen Belastungsstörung, einer Zwangsstörung, einer Persönlichkeitsstörung oder einer klinischen Angststörung aufweisen. Weitere Ausschlusskriterien waren Substanzabhängigkeit, eine Hirnverletzung in der Vorgeschichte, ein früherer Bewusstseinsverlust von mehr als zehn Minuten sowie jegliche Kontraindikationen für eine MRT. Die Teilnehmer füllten den Web-Q-Fragebogen zur Beurteilung der psychischen und physischen Gesundheit sowie den SPHERE-12 (Somatic and Psychological Health Report) und die DASS (Depression and Anxiety Stress Scale) aus. Die Probanden durften keine körperlichen Beeinträchtigungen haben, die ihre Fähigkeit, die Testbatterie auszufüllen, beeinträchtigen könnten, und mussten geistig in der Lage sein, die Studienverfahren und Anweisungen zu verstehen und zu befolgen.
Die Teilnahme der Probanden wurde von der örtlichen Prüfungskommission (Institutional Review Board), der Ethikkommission der Universität von Sydney, genehmigt. Nachdem die Studienverfahren in Übereinstimmung mit den ethischen Richtlinien der Prüfungskommission ausführlich erläutert worden waren, gaben die Teilnehmer ihre schriftliche Einwilligung. Bei minderjährigen Teilnehmern unter 18 Jahren wurde die Zustimmung zur Teilnahme durch einen Erziehungsberechtigten erteilt.
Kognitive Testbatterie
In dieser Studie wurde eine computergestützte kognitive Testbatterie, „WebNeuro“ [Silverstein et al., 2007], verwendet. WebNeuro besteht aus 12 Tests, deren Durchführung 30-40 Minuten dauert. Die Teilnehmer saßen vor einem Computer in einem schall- und lichtgedämpften Raum mit einer Umgebungstemperatur von ca. 24 Grad Celsius. Die Testanweisungen wurden unmittelbar vor der Testdurchführung und den Testmaterialien auf dem Bildschirm angezeigt. Ein Prüfer gab Hilfestellung ( ), wenn dies erforderlich war, insbesondere bei jüngeren Teilnehmern, zusätzlich zu einem interaktiven, automatisierten Protokoll, das das Verständnis des Tests sicherstellte. Wenn der Teilnehmer drei Übungsversuche für einen Test nicht bestand, wurde er automatisch zum nächsten Test weitergeleitet.
Dieselbe Testbatterie läuft auch in einem speziellen Touchscreen-Format, bekannt als „IntegNeuro“ [Clark et al., 2006]. Die Reproduzierbarkeit der computergestützten Plattformen WebNeuro und IntegNeuro wurde nachgewiesen [Silverstein et al., 2007]. Diese kognitiven Tests (im Touchscreen-Format) haben nachweislich eine ausgezeichnete Validität in Bezug auf die Beziehungen zu herkömmlichen Papier-und-Bleistift-Messungen ähnlicher Konstrukte kognitiver Leistungen [Paul et al., 2005; Silverstein et al., 2007]. Die Tests haben auch eine solide Test-Retest-Reliabilität [Williams et al., 2005] und eine solide standortübergreifende Zuverlässigkeit [Paul et al., 2007]. WebNeuro-Alters- und Geschlechtsnormen über 9 Jahrzehnte wurden für eine Normstichprobe von 1000 Teilnehmern erstellt [Mathersul et al., 2009; Williams et al., 2009]. Auf der Grundlage dieser Normen wird die Leistung jedes Studienteilnehmers bei jedem einzelnen Aspekt jedes Tests (Reaktionszeit, Genauigkeit usw.) als z-Score relativ zum normativen Mittelwert von 0 ausgedrückt, wobei positive z-Scores eine überdurchschnittliche Leistung und negative Werte eine unterdurchschnittliche Leistung widerspiegeln. Die Gesamtwerte werden dann als Durchschnitt der einzelnen Testergebnisse für jeden Test berechnet, wie in Tabelle 1 zusammengefasst. Diese zusammenfassenden Werte wurden entsprechend dem Konstruktbereich, den sie repräsentieren, gekennzeichnet.
Tabelle 1. Zusammenfassung der Aufgaben der WebNeuro-Testbatterie – die Konstrukte, die sie bewerten, die einzelnen Testergebnisse, die sie erzeugen, und der Bereich der zusammenfassenden Bewertung
| Aufgabe Name | Beschreibung | Funktion | Variablen | Bereich |
| Auswahl Reaktionszeit | Die Teilnehmer schauen auf den Computerbildschirm, während einer Reihe von Versuchen einer von vier Zielkreisen in pseudozufälliger Reihenfolge aufleuchtet. Bei jedem Versuch muss der Teilnehmer die Maus benutzen und so schnell wie möglich auf den leuchtenden Kreis klicken. Es werden 20 Versuche mit einer zufälligen Verzögerung zwischen den Versuchen von 2-4 Sekunden durchgeführt. | Einfache Entscheidungsfindung | Mittlere Reaktionszeit | Effizienz der Informationsverarbeitung |
| Ziffernspanne (vorwärts) | Den Teilnehmern wird auf dem Computerbildschirm eine Reihe von Ziffern im Abstand von einer Sekunde gezeigt. Der Teilnehmer muss die Ziffern in Vorwärtsreihenfolge abrufen, indem er sie mit der Maus eingibt. | Arbeitsgedächtnis | Abrufspanne | Gedächtnis |
| Kontinuierlicher Leistungstest | Eine Reihe von ähnlich aussehenden Buchstaben (B, C, D oder G) wird dem Teilnehmer auf dem Computerbildschirm für 200msec im Abstand von 2,5 sec präsentiert. Wenn derselbe Buchstabe zweimal hintereinander erscheint, wird der Teilnehmer aufgefordert, die Leertaste zu drücken. | Anhaltende Aufmerksamkeit | Mittlere Reaktionszeit | Aufmerksamkeit |
| Falsch verpasste Fehler | ||||
| Falscher Alarm Fehler | ||||
| Umschalten der Aufmerksamkeit | Die Teilnehmer müssen 13 Ziffern (1-13) und 12 Buchstaben (A-L) in aufsteigender Reihenfolge von abwechselnden Ziffern und Buchstaben identifizieren (z. B. 1 A 2 B 3 C …). Dies erfordert, dass der Teilnehmer seine Aufmerksamkeit zwischen mentalen Aufgaben (Buchstaben- und Zahlenreihenfolge) umschalten muss, wobei Konstrukte bewertet werden, die denen entsprechen, die mit Trail Making B (Reitan, 1958) bewertet werden. | Kognitive Flexibilität | Zeit zur Vervollständigung | Effizienz der Informationsverarbeitung |
| Anzahl der Fehler | ||||
| Labyrinth | Ein Test zur Bewältigung eines Labyrinths, bei dem Planung, Fehlerkontrolle und Entscheidungsfindung getestet werden. Variante des Austin-Labyrinths. Die Teilnehmer navigieren mit den Pfeiltasten auf der Tastatur durch das Gitter. | Exekutive Funktion | Zeit der Beendigung | Bereich der Exekutivfunktion |
| Anzahl der Fehler | ||||
| Go/No-Go | Die Teilnehmer reagieren (mit der Leertaste) so schnell wie möglich auf das Wort „drücken“ in GRÜN (um das automatische Reagieren zu bewerten) und hemmen diese Reaktionen sofort, wenn „drücken“ in ROT angezeigt wird. | Unterdrückung | Variabilität der Reaktionszeit | Impulsivität |
| Gesamtfehler | ||||
| Gedächtnis Wiedererkennen | Den Teilnehmern werden 12 englische Wörter vorgelegt, die sie sich einprägen und später aus dem Gedächtnis erkennen sollen. Etwa 10 Minuten später, nach einer Reihe von Zwischenaufgaben, wird ein verzögerter Gedächtniserkennungsversuch durchgeführt. | Gedächtnis | Sofortiges Erinnern insgesamt | Gedächtnis |
| Verzögertes Erinnern | ||||
| Test zur Identifizierung von Emotionen | Den Teilnehmern wird eine Reihe von Gesichtern präsentiert, die verschiedene Emotionen ausdrücken. Die Teilnehmer müssen sofort erkennen, welche Emotion das jeweilige Gesicht ausdrückt, indem sie mit der Maus die Beschriftung am unteren Rand des Computerbildschirms auswählen. | Erkennung von Emotionen | Reaktionszeit für Angst, Wut und traurige Gesichter | Identifizierung von Emotionen |
| Verzögertes Emotionspriming | Nach einer Pause von 20 Minuten werden die Teilnehmer gebeten, die Gesichter zu erkennen, die in der vorherigen Aufgabe zur Identifizierung von Emotionen präsentiert wurden. Auf diese Weise werden Verzerrungen bei der Erkennung festgestellt, die durch den indirekten Effekt der dargestellten Gesichtsemotionen hervorgerufen werden. | Implizites emotionales Priming | Reaktionszeit für angewiderte und glückliche Gesichter | Emotionale Verzerrung |
Bild-Akquisition
Die MR-Bildgebung für diese Kohorte wurde mit einer 8-Kanal-Phased-Array-Kopfspule an einem 3T GE Signa Twinspeed HDxT MR-Scanner (GE Healthcare, Milwaukee, Wisconsin) in der Abteilung für Radiologie des Westmead Hospital durchgeführt. T1-gewichtete Bilder wurden in der Sagittalebene mit einer 3D SPGR-Sequenz aufgenommen (FOV = 256 mm, TR = 8,3 ms; TE = 3,2 ms; Flip Angle = 11°; TI = 500 ms; NEX = 1 und ASSET = 1,5; Frequenzrichtung: S/I). Insgesamt wurden 180 zusammenhängende 1 mm-Schichten mit einer 256 × 256-Matrix und einer Auflösung in der Ebene von 1 mm × 1 mm aufgenommen, was zu isotropen Voxeln in einer Gesamtscanzeit von 7 Minuten und 12 Sekunden führte.
Voxel-basierte Morphometrie-Analyse/Struktur-MRT-Datenanalyse
Strukturelle Hirnbilder wurden vor der Verarbeitung visuell auf Qualitätskontrolle und Artefakterkennung geprüft. Die Bilder wurden mit der VBM8-Toolbox (http://dbm.neuro.uni-jena.de/vbm.htmal) und dem SPM8-Softwarepaket (http://www.fil.ion.ucl.ac.uk/spm) verarbeitet, die auf MATLAB R2010a (Mathworks) laufen. Für alle Optionen wurden Standardparameter verwendet. Für Querschnittsanalysen wurden die Daten von Zeitpunkt 1 segmentiert und mit Hilfe von hochdimensionalem DARTEL auf den Standard-MNI-Raum normalisiert, und es wurde eine nichtlineare Modulation angewendet. Durch diese Modulation wird sichergestellt, dass die normalisierten Bilder das absolute Volumen der grauen Substanz, korrigiert um die Gesamtgröße des Gehirns, beibehalten.
Für die Längsschnittanalyse wurde die Vorverarbeitung mit dem standardmäßigen Längsschnitt-Preprocessing-Stream der VBM8-Toolbox durchgeführt. Kurz gesagt, werden die Scans zum Zeitpunkt 2 (T2) mit den Scans zum Zeitpunkt 1 (T1) für jeden Probanden getrennt registriert, um ein Durchschnittsbild zu erstellen. Die T1- und T2-Bilder werden dann neu ausgerichtet und auf Signalinhomogenität in Bezug auf das mittlere Bild korrigiert. T1, T2 und das mittlere Bild werden dann in graue Substanz, weiße Substanz und Hirn-Rückenmarks-Flüssigkeit segmentiert und anhand der Segmentierungen des mittleren Bildes normalisiert (SPM8-Standardvorlage). Da die Auswirkung der Modulation auf longitudinale Daten eher subtil ist und Vergleiche innerhalb eines Subjekts stattfinden, wird die Modulation nicht im longitudinalen Verarbeitungsmodul der Toolbox durchgeführt. Für beide Analysen wurden die Bilder visuell auf korrekte Registrierung und Segmentierung geprüft, und die Homogenität der Stichprobe wurde anhand der Kovarianz überprüft. Alle Bilder oberhalb oder unterhalb von 2SDs der Stichprobe wurden überprüft. Die meisten Fehler waren auf Fehler bei der Gehirnextraktion zurückzuführen. In diesen Fällen wurde die Gehirnextraktion mit dem Gehirnextraktionstool in FSL durchgeführt und die Daten wurden erneut verarbeitet. Schließlich wurden die neu ausgerichteten und normalisierten Graue-Materie-Segmente mit einem 8 mm FWHM Gauß-Kernel geglättet.
Zweitens führten wir eine ROI-Analyse durch, um die strukturellen Veränderungen in bestimmten Regionen des CCN zu analysieren. Die CCN-Knoten (dACC, DLPFC und DPC) wurden auf der Grundlage einer kürzlich durchgeführten Meta-Analyse funktioneller Neuroimaging-Studien mit einer Reihe von kognitiven Aufgaben definiert [Niendam et al, 2012]: (1) Die DLPFC-Aktivierung umfasste Knoten im mittleren frontalen Gyrus (L: -40, 26, 28 und R: 40, 30, 28) und im inferioren frontalen Gyrus (L: -42, 4, 30 und R: 44, 6, 32); (2) die DPC-Aktivierung umfasste Knoten im oberen parietalen Lobulus (L: -28, -60, 44 und R: 28, -60, 44), im unteren Scheitellappen (L: -38, -52, 40 und R: 38, -50, 42) und im Precuneus (L: -6, -62, 44 und R: 8, -68, 46) und (3) die dACC-Aktivierung befand sich im Gyrus cinguli (L: -2, 16, 40 und R: 2, 16, 40) (siehe Abb. 2 für die visuelle Darstellung). 2 für die visuelle Darstellung). Unter Verwendung dieser Koordinaten erstellten wir sphärische ROIs mit einem Radius von 8 mm und extrahierten die Rohvolumina zu beiden Zeitpunkten mit dem Marsbar-Tool (http://marsbar.sourceforge.net/).
Abbildung 2 hier in PowerPoint
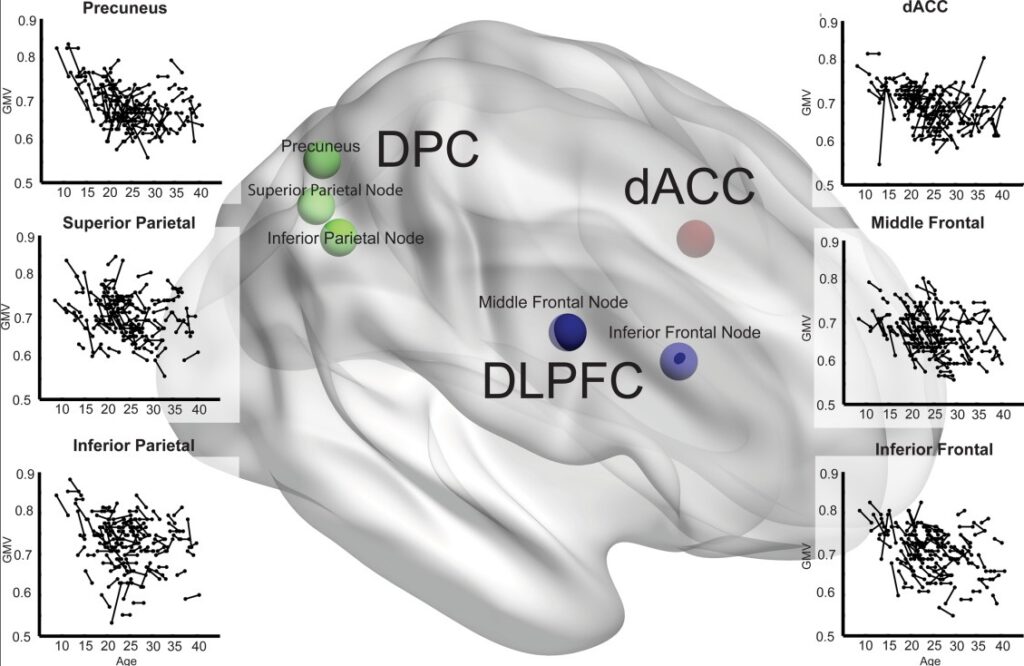
Veränderung des Volumens der grauen Substanz im kognitiven Kontrollnetzwerk mit dem Alter. Die Längsverläufe des Volumens der grauen Substanz (GMV) in den rechten DLPFC-, DPC- und dorsalen anterioren cingulären Cortex- (dACC-) Knoten des CCN sind als Streudiagramme des Volumens der grauen Substanz (alle y-Achsen) und des Alters (alle x-Achsen) dargestellt. Jeder Scan wird durch einen Punkt dargestellt, und wiederholte Scans sind durch Linien verbunden. Die Koordinaten der CCN-Knoten sind in einem 3D-Gehirn eingezeichnet: DLPFC (blau), einschließlich der Knoten inferior frontal (BA9) und middle frontal (BA46), dACC (rot) und DPC (grün), einschließlich der Knoten superior (BA7), inferior (BA40) und precuneus (BA7). [Die Farbabbildung kann unter wileyonlinelibrary.com eingesehen werden.]
Statistische Auswertungen
Die Rohvolumina der grauen Substanz und die z-Scores für die kognitiven Aufgaben wurden auf Normalität getestet und gegen das Alter aufgetragen, um visuell auf Ausreißer zu prüfen. Zur Bewertung der Querschnittsbeziehungen zwischen den Ausgangsvolumina oder den Ergebnissen der kognitiven Bereiche und dem Alter wurde ein allgemeines lineares Modell verwendet, bei dem das Geschlecht und die Wechselwirkungen zwischen Alter und Geschlecht berücksichtigt wurden. Da in früheren Studien eine nichtlineare Veränderung der grauen Substanz beobachtet wurde, wurde dieses Maß unter Einbeziehung des Alters2erneut getestet, um etwaige quadratische Beziehungen zu ermitteln.
Um zu prüfen, ob diese Querschnittsbeziehungen mit dem Alter im Laufe der Zeit beibehalten werden, wurde die Pearson-Korrelation zwischen der Veränderung der Werte oder der Veränderung des Volumens und dem Alter (bei T1) anhand von Längsschnittdaten berechnet. Die Veränderung des Volumens der grauen Substanz (ΔGMV) und der kognitiven Z-Scores (ΔZ-Score) wurden berechnet, indem das Volumen der grauen Substanz (oder der Score) zum Zeitpunkt 1 (T1) vom Zeitpunkt 2 (T2) subtrahiert und durch die genaue Anzahl der Jahre zwischen den auf zwei Jahre angepassten Scans dividiert wurde. Wie unten:
Die Zeit zwischen den Scans wurde auf zwei Jahre angepasst, da die durchschnittliche Zeit zwischen den Untersuchungen in der Kohorte 1,94 Jahre betrug.
Wir testeten auch die Querschnittsbeziehungen zwischen der kognitiven Leistung und der Struktur der grauen Substanz unter Verwendung des allgemeinen linearen Modells, das für das Geschlecht und das Alter als Kovariate kontrolliert. Um dies im Längsschnitt zu testen, wurden partielle Korrelationen unter Kontrolle des Alters und des Geschlechts durchgeführt, um die Beziehung zwischen der ΔGMV in allen Knotenpunkten und dem ΔZ-Score für jeden kognitiven Bereich zu schätzen.
Schließlich wurden partielle Korrelationen unter Kontrolle von Alter und Geschlecht durchgeführt, um die Beziehung zwischen den Veränderungen des Volumens der grauen Substanz in den zwölf Knoten des CCN zu untersuchen.
Für alle Analysen wurde die Bonferroni-Korrektur für Mehrfachvergleiche auf ein nicht bereinigtes Signifikanzniveau von P < 0,05 angewendet. Da wir insgesamt zwölf Knoten des CCN (sechs rechts, sechs links) analysierten, wurde ein Wert von P < 0,004 (0,05/12) als signifikant angesehen. Wir geben auch Ergebnisse auf Trennebene an, die die Korrektur für Mehrfachvergleiche nicht überlebt haben (0,004 < P < 0,05).
ERGEBNISSE
Demografische Daten
Die demografischen Merkmale sowohl der Querschnitt- als auch der Längsschnittkohorte sind in Tabelle 2 zusammengefasst. Die Altersverteilung der Teilnehmer an der Längsschnittstudie ist in Abbildung 1 zu sehen. Um festzustellen, ob die psychische Gesundheit der Teilnehmer über beide Zeitpunkte hinweg stabil war, wurden die mit dem DASS42-Fragebogen für Depression, Angst und Stress erhobenen Daten verwendet. Die Querschnittskohorte hatte einen Durchschnittswert von 3,2 ± 4,1 (Depression), 2,6 ± 3,1 (Angst) und 6,0 ± 5,1 (Stress), was für die Bevölkerung im normalen Bereich liegt. In der Längsschnittgruppe hatte die Stichprobe zum Zeitpunkt 1 eine durchschnittliche Punktzahl von 3,4 ± 4,1 (Depression), 2,9 ± 3,3 (Angst), 6,12 ± 5,0 (Stress) und zum Zeitpunkt 2 durchschnittliche DASS-Werte von 4,3 ± 5,1, 2,5 ± 3,1, 6,5 ± 5,4 für jede der Messungen. Die Analyse mit einem T-Test für gepaarte Stichproben zeigte, dass die DASS-Werte zum Zeitpunkt 1 (T1) und zum Zeitpunkt 2 (T2) nicht signifikant unterschiedlich waren. In allen drei Gruppen fielen über 90 % der Stichprobe in den normalen bis leichten Bereich der DASS-Messungen.
Tabelle 2. Demografische Daten der Teilnehmer
| Querschnittliche Kohorte | Längskohorte | ||
| Zeitpunkt 1 (n = 176) | Zeitpunkt 1 (n = 111) | Zeitpunkt 2 (n = 111) | |
| Alter, y, Mittelwert ± SD (Bereich) | 24 ± 7 (8-38) | 24 ± 7 (8-38) | 26 ± 7 (10-40) |
| Geschlecht, % männlich | 53 | 55 | – |
| Ausbildungsjahre, Median (Spanne) | 16 (2-18) | 16 (2-18) | 16 (2-18) |
| Rechtshändigkeit, % rechts | 86 | 87 | – |
| DASS42- Depressionswert, Mittelwert±SD | 3.18 ± 4.09 | 3.60 ± 4.39 | 4.22 ± 4.93 |
| DASS42- Angstwert, Mittelwert ±SD | 2.61 ± 3.04 | 2.97 ± 3.28) | 2.64 ± 3.12 |
| DASS42-Stress-Score, Mittelwert ±SD | 6.01 ± 5.06) | 6.30 ± 5.02 | 6.59 ± 5.44 |
Querschnittliche Zusammenhänge
Alter und Volumen der grauen Substanz
Im Querschnitt wurde festgestellt, dass das Alter eine signifikante negative Beziehung (Bonferroni, P < 0,004) zu allen Knoten des CCN (P < 0,002) aufweist, mit Ausnahme des linken mittleren frontalen und inferioren frontalen DLPFC (P = 0,012; P = 0,008), die die Korrektur für multiple Vergleiche nicht überstanden. In unserer Kohorte waren keine Wechselwirkungen zwischen Alter und Geschlecht signifikant. Eine Zusammenfassung aller Beziehungen zwischen Alter, Volumen der grauen Substanz und Verhalten findet sich in Tabelle 3.
Tabelle 3. Zusammenfassung der Ergebnisse
| Querschnittliche Assoziationen zwischen dem Volumen der grauen Substanz und | Veränderung des Volumens der grauen Substanz im Längsschnitt Korrelationen mit | |||||||||
| Alter | Geschlecht | Verhalten | Alter | Δ Verhalten | ||||||
| Region | Knoten | Koeffizienta | P-Werte | P-Werte | Koeffizientb | P-Werte (Aufgabe) | Korr.c | P-Werte | Korr.d | P-Werte (Aufgabe) |
| dACC | Links | -0.004 | <0.001** | NS | – | NS | – | NS | -0.240 | 0,030 (Informationsverarbeitung) |
| Rechts | -0.004 | <0.001** | 0,028 (F>M) | – | NS | – | NS | -0.278 | 0,011 (Informationsverarbeitung) | |
| DLPFC | Links inferior frontal | -0.003 | 0.008 | NS | 0.021 | 0,015 (Exekutivfunktion) | – | NS | -0.282 | 0,01 (Exekutivfunktion) |
| Rechts inferior frontal | -0.004 | 0.001** | NS | – | NS | – | NS | – | NS | |
| Linke mittlere Frontpartie | -0.003 | 0.012 | NS | 0.047 | 0,001** (Exekutivfunktion) | – | NS | -0.233, -0.252 | 0,036 (Exekutivfunktion), 0,022 (Emotions-ID) | |
| Rechts Mitte frontal | -0.003 | 0.002** | NS | – | NS | 0.234 | 0.013 | – | NS | |
| DPC | Links inferior parietal | -0.005 | <0.001** | NS | – | NS | – | NS | – | NS |
| Rechts inferior parietal | -0.005 | <0.001** | NS | – | NS | – | NS | -0.339 | 0,002** (Exekutivfunktion) | |
| Links superior parietal | -0.005 | <0.001** | NS | 0.020 | 0,01 (Exekutivfunktion) | – | NS | -0.302 | 0,006 (Exekutivfunktion) | |
| Rechts Superior Parietal | -0.003 | 0.001** | NS | – | NS | – | NS | -0.355 | 0,001** (Exekutivfunktion) | |
| Linker Präkuneus# | -0.025 | <0.001** | 0,033 (F>M) | – | NS | – | NS | – | NS | |
| Rechter Präkuneus# | -0.022 | <0.001** | NS | – | NS | – | NS | -0.249 | 0,024 (Informationsverarbeitung) | |
- #Zeigte quadratische Zusammenhänge mit dem Alter.
- **Signifikant auf korrigiertem Niveau (Bonferroni, P < 0,004).
- NS-nicht signifikant.
- aAlters- und Geschlechtseffekte mit GMV wurden innerhalb desselben allgemeinen linearen Modells geschätzt.
- bAllgemeines lineares Modell zur Schätzung des Zusammenhangs zwischen jeder Verhaltensmaßnahme und GMV, Alter nicht kontrolliert.
- cPearsonsche Korrelation.
- dBivariate Korrelation, kontrolliert für Alter und Geschlecht.
Alter und kognitive Verhaltensmessungen
Es gab keine signifikanten Zusammenhänge zwischen den kognitiven Werten und dem Alter und keine Auswirkungen auf das Geschlecht oder Alter*Geschlecht für einen der kognitiven Bereiche.
Volumen der grauen Substanz und kognitive Verhaltensmaße
Es bestand ein signifikanter Zusammenhang zwischen dem Volumen der grauen Substanz im linken mittleren frontalen Knoten (P = 0,001) und auch Trends für den linken inferioren frontalen Knoten (P = 0,015) des DLPFC und den linken superioren parietalen Knoten des DPC (P = 0,01) mit der Exekutivfunktion. Diese Beziehungen blieben jedoch nicht bestehen, wenn das Alter berücksichtigt wurde.
Longitudinale Beziehungen
Korrelation der Veränderung des Volumens der grauen Substanz mit dem Alter
Abbildung 2 zeigt die individuellen Volumina der rechten Knoten des CCN bei T1 und T2 im Vergleich zum Alter. Nur der rechte mittlere frontale Knoten des DLPFC wies eine tendenzielle Beziehung zum Alter auf, wobei sich das Volumen mit zunehmendem Alter weniger veränderte (P = 0,013). Keiner der anderen CCN-Knoten zeigte eine signifikante Beziehung zwischen Alter und Veränderung des Volumens der grauen Substanz im Längsschnitt.
Korrelation der Veränderung von kognitiven Verhaltensmaßen mit dem Alter
Es schien in keinem Bereich eine Korrelation zwischen der Veränderung der kognitiven Leistung und dem Alter zu geben.
Korrelation der Veränderung des Volumens der grauen Substanz mit der Veränderung der Verhaltensleistung
Die Leistung der Exekutivfunktionen korrelierte signifikant mit dem Volumen der grauen Substanz des rechten inferioren (P = 0,002) und superioren parietalen Lappens (P = 0,001) der DPC. Es wurden auch Trends mit einer negativen Korrelation im linken inferioren Frontallappen (P = 0,010) und im linken mittleren Frontallappen (P = 0,036) im DLPFC und im linken superioren parietalen Knoten des DPC (P = 0,006) beobachtet (siehe Abb. 3A). Darüber hinaus zeigten sich Tendenzen einer negativen Korrelation zwischen den Ergebnissen der Informationsverarbeitungseffizienz und dem Volumen der grauen Substanz im bilateralen dACC (L: P = 0,03, R: 0,011) und im rechten Precuneus im DPC (P = 0,024) (siehe Abb. 3B) sowie für die Emotionsidentifikationsleistung und das Volumen der grauen Substanz im linken mittleren frontalen Lobulus des DLPFC (P = 0,022). Für alle Regionen wurde eine Abnahme des Volumens bei Verbesserung der kognitiven Leistung beobachtet.
Abbildung 3 hier in PowerPoint
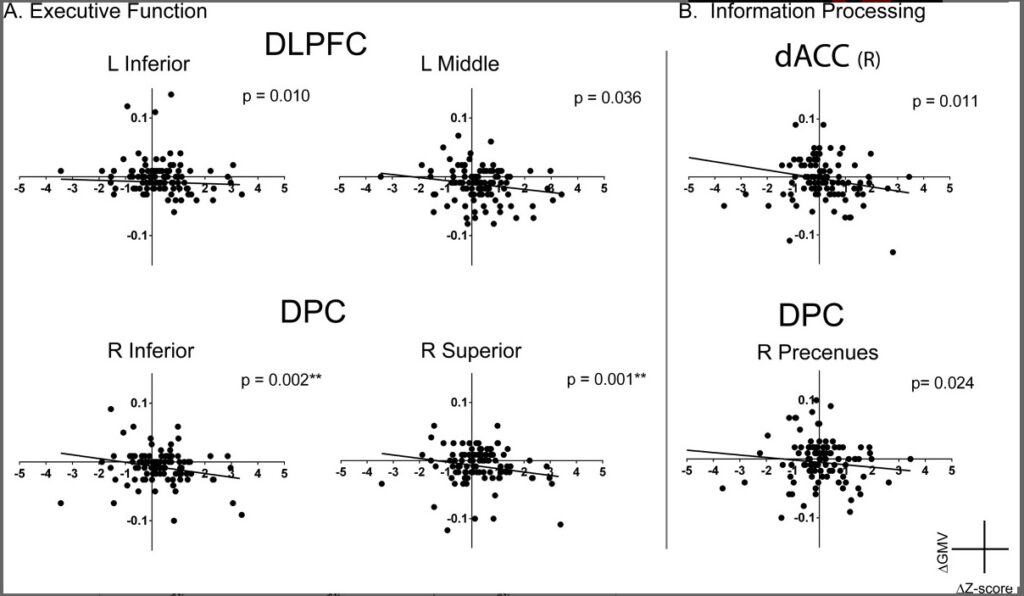
Zusammenhang zwischen der Veränderung des Volumens der grauen Substanz und der kognitiven Leistung. Die Beziehung zwischen der Veränderung des Volumens der grauen Substanz (ΔGMV) und der Veränderung der kognitiven Werte (ΔZ-Score) unter Kontrolle von Alter und Geschlecht zeigt a) die Veränderungen des Volumens der grauen Substanz im dorsolateralen präfrontalen Kortex (DLPFC) und im dorsalen parietalen Kortex (DPC), die mit einer Verbesserung der exekutiven Funktion einhergehen, und b) die Veränderungen des Volumens der grauen Substanz im dorsalen anterioren cingulären Kortex (dACC) und im DPC, die mit einer verbesserten Informationsverarbeitung einhergehen. Bei beiden Aufgaben ist eine stärkere Abnahme des Volumens der grauen Substanz über zwei Jahre signifikant mit einer Verbesserung der kognitiven Leistung verbunden.
Gibt es eine kodierte anatomische Veränderung im CCN?
Partielle Korrelationsanalysen, bei denen Alter und Geschlecht kontrolliert wurden, ergaben hochsignifikante Ergebnisse (P < 0,0001) zwischen den bilateralen dACC-Knoten, den bilateralen inferioren frontalen und den linken mittleren frontalen Knoten des DLPFC sowie den bilateralen inferioren und superioren parietalen Knoten des DPC. Der linke Precuneus wies eine signifikante Korrelation (P < 0,0031) mit dem bilateralen Cingulum, den bilateralen inferioren frontalen Knoten, dem linken mittleren frontalen Knoten, den bilateralen inferioren parietalen Knoten und den bilateralen superioren parietalen Knoten sowie mit dem rechten Precuneus auf. Der rechte Precuneus wies eine signifikante Korrelation (P < 0,001) mit allen außer dem linken inferioren und dem bilateralen mittleren frontalen DLPFC-Knoten auf. Am wenigsten korreliert mit allen anderen Bereichen im CCN war der rechte mittlere frontale Knoten des CCN, der nur mit den rechten inferioren und linken mittleren frontalen DLPFC-Knoten signifikant (P < 0,003) korreliert war.
DISKUSSION
Diese Studie liefert den ersten direkten Beweis dafür, dass die strukturelle Reifung von Regionen des CCN die Entwicklung exekutiver Funktionen unterstützt. Indem wir in einer Längsschnittstudie kognitive Leistungen und strukturelle Hirnveränderungen im Laufe der Zeit abbildeten, beobachteten wir nicht nur koordinierte Veränderungen der grauen Substanz in diesem Netzwerk, sondern auch eine signifikante Assoziation dieser Veränderungen mit einer Verbesserung der kognitiven Fähigkeiten während einer Schlüsselphase der Neuroentwicklung. Wir fanden heraus, dass unabhängig vom Alter eine Abnahme des Volumens der grauen Substanz im posterioren parietalen und dorsolateralen präfrontalen Kortex mit einer verbesserten Leistung im Bereich der exekutiven Funktionen korreliert war. Darüber hinaus war eine Abnahme des Volumens der grauen Substanz im bilateralen cingulären Kortex und im rechten Precuneus mit einer verbesserten Leistung im Bereich der Informationsverarbeitung und im linken dorsolateralen präfrontalen Kortex bei der Emotionsverarbeitung verbunden (beide auf Trendniveau). Etwas überraschend war, dass die longitudinale Veränderung sowohl des Volumens der grauen Substanz im CCN als auch der kognitiven Leistung in dieser Stichprobe nicht mit dem Alter zusammenhing.
Es wurde beobachtet, dass sich die Struktur der grauen Substanz im menschlichen Gehirn während der gesamten Lebensspanne verändert. Querschnittsbefunde haben einen nichtlinearen Rückgang der grauen Substanz vom Alter von etwa 7 bis zum Alter von 60 Jahren beschrieben [Grieve et al., 2005, 2011; Pfefferbaum et al., 1994; Sowell et al., 2003]. Während die bisherigen Längsschnittstudien einen Rückgang der grauen Substanz während der Adoleszenz bis in die Zwanzigerjahre belegen, gibt es nur wenige Längsschnittdaten zu Veränderungen der grauen Substanz im zweiten, dritten und vierten Lebensjahrzehnt [Gogtay et al., 2004; Lenroot et al., 2007]. Zwei frühere Längsschnittstudien mit großen Kohorten, die Probanden im Alter vom ersten bis zum dritten Lebensjahrzehnt umfassten, beschreiben eine Periode der anfänglichen Zunahme in der Kindheit, gefolgt von einem Rückgang in der Adoleszenz und einer anschließenden Stabilisierung der Veränderungen der grauen Substanz [Lenroot et al., 2007; Shaw et al., 2008]. Darüber hinaus wurden auch geschlechtsspezifische Unterschiede in den Reifungstendenzen festgestellt [Lenroot et al., 2007]. Dieses Muster der Abnahme der grauen Substanz im Querschnitt mit dem Alter wurde auch in unserer Kohorte beobachtet, allerdings verlief der Trend meist linear. Im Gegensatz dazu zeigen die beobachteten Längsschnittbefunde, dass die Veränderungsrate des Volumens der grauen Substanz weitgehend unabhängig vom Alter war, mit Ausnahme eines moderaten Trends im rechten mittleren frontalen Knoten des DLPFC. Eine mögliche Erklärung für diese Unterschiede ist der hohe Anteil der Teilnehmer in den 20er und 30er Jahren, insbesondere im Vergleich zu den unter 18-Jährigen, die ein Mindestalter von acht Jahren hatten. Dieser überproportionale Anteil an jüngeren Teilnehmern könnte die Aussagekraft der Daten beeinträchtigt haben. Dies ist besonders relevant, wenn man die Daten mit denen der beiden zuvor erwähnten Längsschnittstudien vergleicht, die eine überwiegend jüngere Kohorte mit einem Mindestalter von drei Jahren hatten. Dies könnte auch dazu geführt haben, dass geschlechtsspezifische Effekte, von denen man annimmt, dass sie altersabhängig sind und die Trajektorien der grauen Substanz im gesamten Kortex beeinflussen [Lenroot et al., 2007], nicht berücksichtigt wurden. Ein weiterer wahrscheinlicher Grund ist, dass unsere Ergebnisse auf Daten für nur zwei Zeitpunkte basieren, während die Studien von Lenroot et al. und Shaw et al. bei über 30-40 % ihrer großen Stichproben (387 bzw. 375) MRT-Daten von 3 oder 4 Zeitpunkten enthielten, die einen Entwicklungsverlauf über die Zeit besser erfassen [Lenroot et al., 2007; Shaw et al., 2008]. Darüber hinaus konzentrierte sich unsere Studie nur auf Regionen der grauen Substanz innerhalb des CCN, also auf Hirnregionen, die nachweislich im Laufe des Lebens die größte Plastizität aufweisen [Toga et al., 2006].
Es wurde bereits beobachtet, dass strukturelle Veränderungen im Gehirn weitgehend parallel zu kognitiven Meilensteinen verlaufen [Casey et al., 2005]. In dieser Studie haben wir diese Beziehungen in sieben kognitiven Bereichen untersucht. Wir fanden eine Beziehung zwischen den Volumina von DPC und DLPFC und den exekutiven Funktionen sowie zwischen DPC und dACC und der Informationsverarbeitung. Der Bereich der exekutiven Funktion erfasst eine allgemeine Leistung von kognitiven Kontrollfunktionen. Die Informationsverarbeitung greift auch auf ein breiteres Spektrum spezifischer Aufmerksamkeitskontrollfunktionen zu, die zuvor mit der Konnektivität des dACC und des parietalen Kortex in Verbindung gebracht wurden [Mesulam, 1983]. Unsere Ergebnisse unterstützen frühere Längsschnittbefunde aus zwei Studien, von denen eine eine Verbesserung der verbalen Intelligenz in Verbindung mit einer Abnahme der grauen Substanz in frontalen Bereichen sah [Sowell et al., 2004] und die zweite eine Verbesserung des Arbeitsgedächtnisses in Verbindung mit einer Abnahme des Volumens der grauen Substanz im frontalen und parietalen Bereich beobachtete [Tamnes et al., 2013]. Beide Ergebnisse waren auch unabhängig vom Alter [Sowell et al., 2004; Tamnes et al., 2013]. Wir beobachteten auch einen Trend in der Beziehung zwischen DLPFC und Emotionsidentifikation, was die Rolle dieser Region bei der kognitiven Regulation und Verarbeitung von Emotionen unterstützt [Erk et al., 2010]. Trotz der bekannten Rolle einer koordinierten funktionellen Aktivierung des gesamten CCN für eine Reihe von kognitiven Aufgaben fanden wir keine Beziehungen zwischen CCN-Volumina und Bereichen des Gedächtnisses, der Aufmerksamkeit und der Impulsivität oder signifikante Beziehungen für alle Regionen des CCN mit exekutiven Funktionen und der Informationsverarbeitung. Trotz der funktionellen Aktivierung von CCN-Regionen während einer Reihe von kognitiven Aufgaben [Niendam et al., 2012] ist es immer noch möglich, dass diskrete Komponenten und Verbindungen innerhalb des Netzwerks spezifische Verhaltenskorrelate aufrechterhalten und folglich regionalspezifische Strukturänderungen aufweisen.
Darüber hinaus können Unterschiede in der Beziehung zwischen Strukturveränderungen und Verhaltensänderungen auf unterschiedliche Entwicklungsverläufe für bestimmte kognitive Funktionen zurückzuführen sein, da in der Literatur noch kein Konsens darüber besteht, wann die kognitive Kontrolle vollständig ausgereift ist. Studien haben ergeben, dass die im Erwachsenenalter beobachtete kognitive Leistung in der Regel im späten Jugendalter erreicht wird [Bunge et al., 2002]. Es gibt jedoch auch Belege für eine kontinuierliche Verbesserung der kognitiven Leistung bis in die Zwanziger- und Dreißigerjahre [Clark et al., 2006]. Wir konnten keine Verbesserungen der kognitiven Fähigkeiten feststellen, die auf ein höheres Alter zurückzuführen waren. Die von uns beobachteten Zusammenhänge zwischen der Struktur der grauen Substanz und der Kognition waren auch unabhängig vom Alter. Die hier und an anderer Stelle beobachteten Zusammenhänge lassen vermuten, dass die Beziehung zwischen abnehmendem Volumen der grauen Substanz und verbesserter Leistung auf altersunabhängige synaptische Abnutzungsprozesse zurückzuführen ist, die aufgrund individueller Unterschiede bei den genetischen Faktoren oder aufgrund erfahrungsbedingter neuronaler Plastizität auftreten. Es hat sich nämlich gezeigt, dass sowohl genetische als auch umweltbedingte Faktoren sowohl der Gehirnstruktur als auch den kognitiven Fähigkeiten zugrunde liegen [Finkel et al., 2005].
Ein weiterer Beleg für aktivitätsbedingte Mechanismen der Netzwerkverfeinerung und der Verbesserung der kognitiven Kontrolle ist die beobachtete Koordinierung der CCN-Entwicklung, also von Bereichen, die bekanntermaßen funktionell miteinander verbunden sind. Raznahan et al., [2011] berichteten, dass anatomische Veränderungen in verschiedenen Regionen des sich entwickelnden Kortex eine hoch korrelierte, nicht zufällige Struktur aufweisen [Raznahan et al., 2011]. Diese Ergebnisse unterstützen frühere Neuroimaging-Studien, die Belege für aktivitätsabhängige strukturelle Plastizität liefern [Draganski et al., 2004; Hyde et al., 2009]. Dies deutet darauf hin, dass Regionen, die im Laufe der Zeit koordinierte Aktivierungsmuster teilen, während und nach der Entwicklung ähnlicheren aktivitätsbedingten Strukturveränderungen unterliegen als funktionell voneinander unabhängige kortikale Regionen. Ein weiterer interessanter Befund ist, dass der rechte mittelfrontale Knoten des DLPFC ein Ausreißer war, der keine starke Korrelation mit anderen Regionen des CCN aufwies. Der rechte mittelfrontale Knoten wies ein lineares Muster des Rückgangs auf, das mit dem Alter eine tendenzielle Längsbeziehung aufwies. Dies deutet auf eine möglicherweise langwierige Entwicklung dieses Teils des DLPFC hin. Eine mögliche Erklärung hierfür ergibt sich aus früheren funktionellen Studien, die eine altersabhängige Lateralisierung des DLPFC während der Entwicklung festgestellt haben [Bunge et al., 2002; Rubia et al., 2006]. Zum Beispiel rekrutieren Kinder bei Arbeitsgedächtnisaufgaben den rechten DLPFC nicht in demselben Maße wie Jugendliche und Erwachsene [MacDonald et al., 2000; Scherf et al., 2006]. Diese neuroentwicklungspsychologischen Befunde geben Aufschluss darüber, dass die Adoleszenz und die frühen Zwanzigerjahre die Zeit mit dem höchsten Risiko für die Entwicklung psychiatrischer Erkrankungen sind [Paus et al., 2008]. Die altersunabhängige Rolle der strukturellen Veränderungen und die Koordinierung dieser Veränderungen im gesamten Netzwerk deuten darauf hin, dass aktivitätsgesteuerte Prozesse eine Schlüsselrolle bei der Gestaltung struktureller Netzwerke spielen, was sich wiederum auf die kognitive Kontrollleistung auswirkt.
Das Verständnis des normalen Verlaufs der Entwicklung der kognitiven Kontrolle und der Beziehung zu strukturellen Veränderungen im Gehirn ist entscheidend für das Verständnis möglicher Ursachen für die bei verschiedenen psychischen Störungen beobachteten Beeinträchtigungen der kognitiven Kontrolle [Phillips et al., 2008; Williams, 2016]. Das Verständnis psychischer Erkrankungen durch eine Perspektive der neuronalen Schaltkreise ermöglicht es der Forschung, die komplexe und heterogene Natur dieser Störungen besser zu verstehen und zu behandeln. Mithilfe von MRT-Studien lassen sich Muster und Stereotypen in der Dysfunktion der Schaltkreise bei verschiedenen Störungen erkennen [Williams, 2016]. Insbesondere im CCN wurde eine Hypoaktivierung des DLPFC und des dACC als mögliche Ursache für einen „kognitiven Dyskontrolltyp“ bei Depressionen identifiziert, da es nicht gelingt, das Grübeln erfolgreich zu dämpfen und negative Reaktionen zu hemmen [Williams, 2016]. Auch wenn noch weitere Forschungsarbeiten erforderlich sind, haben mehrere Studien ergeben, dass eine verringerte funktionelle Aktivität in Bereichen des CCN zur emotionalen Dysregulation bei bipolaren Störungen beitragen kann [Green et al., 2007; Lagopoulos et al., 2007; Phillips et al., 2008]. Durch die Neudefinition psychischer Störungen anhand neuronaler und nicht symptomatischer Messgrößen können diese Studien nicht nur die Sichtweise auf psychische Erkrankungen verändern, sondern auch zur Entwicklung besserer Diagnoseinstrumente und Behandlungen beitragen. Insbesondere das Verständnis der Mechanismen der Neuroentwicklung der kognitiven Kontrollschaltkreise, die in einem Zeitraum stattfinden, in dem ein erhöhtes Risiko für die Entwicklung psychischer Störungen besteht, kann zu einem besseren Verständnis und einer besseren Behandlung psychiatrischer und neurologischer Erkrankungen führen, bei denen die kognitiven Kontrollschaltkreise betroffen zu sein scheinen.
Die Ergebnisse dieser Studie wurden durch mehrere Einschränkungen beeinträchtigt. Die größte Einschränkung für diese Studie ist die geringe Zahl der Teilnehmer unter 18 Jahren im Vergleich zu den über 18-Jährigen (etwa 15 unter 18 Jahren, 100 sind 18-38). Es ist wahrscheinlich, dass die Entwicklungsveränderungen während dieses Zeitraums in den Ergebnissen unterrepräsentiert sind. Zweitens waren die Längsschnittdaten für diese Studie auf nur zwei Scans beschränkt. Wenn mehr Zeitpunkte pro Individuum gemessen werden, erhöht sich die Aussagekraft für die Modellierung von Längsschnittbefunden und ermöglicht eine differenziertere Analyse wie die gemischte Modellierung [Mills et al., 2014]. Drittens ist bekannt, dass die longitudinale Verarbeitung struktureller MRT-Daten durch die Wahl der Vorverarbeitungsmethodik beeinflusst wird, z. B. durch Verzerrungen aufgrund der für die Ausrichtung anderer Zeitpunkt-Scans ausgewählten Zeitpunktdaten und durch Standard-Raumschablonen [Thomas et al., 2009]. In unserer Studie folgten wir der standardmäßig empfohlenen longitudinalen Pipeline für die VBM-Analyse, die die Daten an Scans zum Zeitpunkt 1 ausrichtet. Wir stellen jedoch fest, dass, obwohl unsere Hauptergebnisse robust waren und unverändert blieben, als die Daten von Zeitpunkt 2 zum Abgleich aller Scans verwendet wurden, zwei unserer Trendergebnisse in der zweiten Analyse nicht mehr signifikant waren. Dies betraf die Korrelation zwischen der Veränderung der Emotionsidentifikation und der Veränderung des linken mittleren frontalen Volumens (der P-Wert änderte sich von 0,022 auf 0,079) und die Korrelation zwischen der Veränderung der Informationsverarbeitung und der Veränderung des rechten Precuneus-Volumens (der P-Wert änderte sich von 0,024 auf 0,365). Die vierte Einschränkung besteht darin, dass das Volumen der grauen Substanz als Messgröße sehr individuellen Schwankungen unterworfen sein kann, die auf Lebensstil und Erfahrung beruhen. Ceccarelli et al. haben beispielsweise gezeigt, dass sich das Volumen der grauen Substanz innerhalb von zwei Wochen ändern kann, wenn die Probanden an einem „kognitiven Training“ teilnehmen [Ceccarelli et al., 2009]. Obwohl unsere Stichprobe so ausgewählt wurde, dass sie nur gesunde Kontrollteilnehmer aus der Allgemeinheit umfasst, ist es praktisch nicht möglich, alle individuellen Faktoren zu berücksichtigen, die das Volumen der grauen Substanz beeinflussen könnten. Obwohl wir uns auf das CCN konzentriert haben, liefern die Messungen des Volumens der grauen Substanz nur ein begrenztes Verständnis der Beziehungen zwischen den verschiedenen Knotenpunkten dieses Netzwerks. Ein tieferes Verständnis dieser Beziehungen kann möglicherweise durch die Verwendung von anspruchsvolleren bildgebenden Messungen der strukturellen Konnektivität (Diffusion Tensor Imaging) und der funktionellen Konnektivität (funktionelle MRT) erlangt werden, um die inter- regionalen Interaktionen innerhalb dieses Netzwerks und ihre Verbindungen zur kognitiven Kontrolle umfassend zu untersuchen. Dies wird der Schwerpunkt unserer zukünftigen Arbeit sein.
Zusammenfassend lässt sich sagen, dass diese Ergebnisse die wenigen bereits vorhandenen Forschungsarbeiten ergänzen, die den Verlauf der strukturellen Längsentwicklung über die Kindheit hinaus bis ins zweite bis vierte Lebensjahrzehnt untersuchen. Diese Daten tragen dazu bei, eine normale Ausgangsbasis für das späte Jugend- und frühe Erwachsenenalter zu schaffen, eine Schlüsselperiode der Entwicklung, in der Menschen eine erhöhte Anfälligkeit für psychiatrische Erkrankungen aufweisen. Diese Ergebnisse können in Zukunft genutzt werden, um Marker für eine bessere Diagnose und ein besseres Verständnis der Auswirkungen dieser Störungen auf die Neuroentwicklung zu untersuchen und um zu zeigen, wie sich verbesserte Behandlungsergebnisse auf die Gehirnstruktur auswirken können. Diese Studie liefert direkte Belege für einen altersunabhängigen Zusammenhang zwischen kognitiver Leistung und dem Volumen der grauen Substanz in Schlüsselbereichen des CCN. Darüber hinaus deutet die Korrelation zwischen strukturellen Veränderungen im Laufe der Zeit im CCN auf eine Koordinierung der anatomischen Veränderungen während der Entwicklung hin, die möglicherweise durch die funktionelle Konnektivität zwischen diesen Bereichen beeinflusst werden oder diese wiederum beeinflussen. Weitere Untersuchungen unter Verwendung eines breiteren Spektrums von Neuroimaging-Messungen werden zu einem besseren Verständnis der Mechanismen beitragen, die dieser wichtigen Beziehung zugrunde liegen.
ETHISCHE GENEHMIGUNG
Alle Verfahren, die in Studien mit menschlichen Teilnehmern durchgeführt wurden, entsprachen den ethischen Standards der institutionellen und/oder nationalen Forschungskommission sowie der Erklärung von Helsinki von 1964 und ihren späteren Änderungen oder vergleichbaren ethischen Standards.
EINWILLIGUNG NACH AUFKLÄRUNG
Von allen an der Studie teilnehmenden Personen wurde eine informierte Zustimmung eingeholt. Bei Personen unter 18 Jahren wurde die informierte Zustimmung eines Elternteils oder Erziehungsberechtigten eingeholt.
VERWEISE
Literaturhinweise PDF herunterladen
Zusätzliche Links Über Wiley Online Bibliothek