
Erläuterungen zu diesem Beitrag
Titelei Originalpublikation Deepl Übersetzung, überarbeitet von A. Eckerle
{kind=link}
Abstract
Einleitung: Die Untersuchung neurostruktureller Marker für intellektuelle Begabung (IG) wird das wissenschaftliche Verständnis der Prozesse förödern, die Kindern zu akademischen Höchstleistungen verhelfen.
Methode: Mit Hilfe der strukturellen und diffusionsgewichteten MRT wurden die regionale Hirnform und die Konnektivität von Kindern verglichen, die entweder einen durchnittlichen bis hohen IQ (TD – typically developing children, 12 Probanden) oder einen IQ von über 145 aufwiesen (IG – intellectual gifted, 18 Probanden). (s. Validierung an großer Stichprobe unter „Schlussfolgerungen“)
Ergebnisse: IG hatte größere subkortikale Strukturen und eine robustere mikrostrukturelle Organisation der weißen Substanz zwischen diesen Strukturen in Regionen, die mit explzitem Gedächtnis assoziiert sind. TD hatte mehr verbundene, größere subkortikale Strukturen in Regionen, die mit dem impliziten Gedächtnis verbunden sind.
Schlussfolgerungen: Es wurde festgestellt, dass die Gedächtnissysteme in den Gehirnen von Kindern mit außergewöhnlichen intellektuellen Fähigkeiten im Vergleich zu Gehirnen von Kindern mit normaler Entwicklung anders dimensioniert und verbunden sind. Diese unterschiedlcihen neurologischen Entwicklungsverläufe deuten auf unterschiedliche Lernstrategien hin. es wird ein Spektrum von Intelligenztypen angenommen, das durch ein unterschiedliches Verhältnis von impliziten und expliziten Systemen begünstigt wird und das anhand eines großen externen Datensatzes validiert wurde.
Einführung
Trotz ihrer außergewöhnlichen Lern- und Testeignung wissen wir erstaunlich wenig darüber, wie sich die Lern- und Gedächtnissysteme in intellektuell „begabten“ Gehirnen entwickeln und funktionieren. Kinder mit außergewöhnlicher Intelligenz verfügen in der Regel über effizientere Gedächtnisfunktionen, größere und komplexer organisierte Wissensbestände und sind in der Lage, komplexere kognitive Strategien anzuwenden. Diese stützen sich auf Gedächtnis- und semantische Strukturen, um Probleme schneller oder in einem früheren Alter zu lösen als Kinder mit typischer Entwicklung (Ali et al., 2003, Athanasakis et al., 2014, Colom et al., 2004, Davidson, 1986). So sind sie in der Lage, Wissen bereichsübergreifend zu verallgemeinern, intuitive Sprünge zu machen (Desco et al., 2011) und spontan selektives Lernen zu nutzen sowie Informationen während des Problemlösens zu vergleichen und zu integrieren (Duncan et al., 2000). „Begabte“ Kinder nutzen komplexere kognitive Strategien, um Probleme schneller oder früher zu lösen als TD-Kinder (Ali et al., 2003, Athanasakis et al., 2014, Colom et al., 2004, Davidson, 1986), und die Grundlage für viele dieser fortgeschrittenen kognitiven Fähigkeiten scheint ihr Gedächtnissystem zu sein. Frühere Studien haben eine altersbedingte globale Zunahme der kortikalen Dicke gezeigt, vor allem im Bereich des exekutiven präfrontalen Kortex; es wurden positive Korrelationen zwischen der Integrität der weißen Substanz, die frontale und parietale Regionen verbindet, bei begabten Kindern festgestellt (Geake & Hansen, 2005, Gerig et al., 2001). Darüber hinaus wurden Belege für eine verstärkte oder abweichende neurologische Entwicklungskurve bei mathematisch begabten Kindern gefunden, deren Gehirne dazu neigen, die frontotemporalen Systeme der rechten Hemisphäre selektiv und erfolgreich zu nutzen, was ihnen außergewöhnlich gute Leistungen ermöglicht (O’Boyle et al., 1990, O’Boyle et al., 1995, O’Boyle et al., 2005, Packard & Knowlton, 2002, Pesenti et al., 2001).
Von den wenigen vorhandenen Studien hat sich jedoch keine auf die Gedächtnissysteme konzentriert. Unser Verständnis der Mechanismen, wie Informationen integriert werden, ist noch unzureichend, wie z. B. die regionale Form und die Konnektivität neuronaler Lernnetzwerke. Zwei parallele Gedächtnissysteme sind bei der Untersuchung der Neuroentwicklung von besonderem Interesse. Implizites Lernen beschreibt das automatische Lernen von sozialen, sprachlichen und prozeduralen Aufgaben, die ohne bewusste Anstrengung bewältigt werden. Dieser Mechanismus ermöglicht es Kindern, grammatikalisch korrekte Sätze zu bilden und soziale Normen, wie z. B. Blickkontakt, ohne explizite Anweisungen zu verstehen (Gonring et al., 2017). Explizites Lernen beinhaltet dagegen eine absichtliche, bewusste Anstrengung, um Informationen zu behalten oder abzurufen, z. B. das Erinnern an Vergangenes oder das Aufsagen von auswendig gelernten Fakten. Implizites Lernen ist in den frühen Stadien der Neuroentwicklung von wesentlicher Bedeutung, während Erwachsene mehr auf das explizite Gedächtnis angewiesen sind, wahrscheinlich weil ein regelbasierter Ansatz (im Unterricht, Eck.) schneller und einfacher zu kommunizieren ist und zu einer Alles-oder-nichts-Beherrschung führt (Gray et al., 2003). In der typischen Entwicklung werden die subkortikalen Strukturen, die das implizite Lernen ermöglichen (z. B. Striatum), im ersten Lebensjahr entwickelt, während die expliziten Gedächtnisstrukturen (z. B. Hippocampus) länger brauchen, um reifen.
Wir haben die Neuroanatomie, die diesen beiden parallelen Gedächtnissystemen, dem expliziten und dem impliziten Gedächtnis, zugrunde liegt, in zwei Stichproben untersucht. Zunächst untersuchten wir Probanden, die speziell wegen ihrer hohen intellektuellen Begabung (IQ >145) rekrutiert wurden, zusammen mit TD-Kindern. Wir haben strukturelle und diffusionsgewichtete MRT- und neuropsychologische Methoden verwendet, um die regionale Hirnform und Konnektivität von 12 Kindern mit durchschnittlichem bis hohem IQ (90-130, Mittelwert= 124± 10,9; Alter= 10,7± 1,86; 58% weiblich) zu vergleichen; und 18 mit sehr hohem IQ (145-170, Mittelwert= 153± 11,4; Alter = 10,2± 2,02; 56 % weiblich). Anschließend haben wir die in dieser Stichprobe entwickelte Heuristik an einer großen, externen Datenbank getestet, um die Ergebnisse zu bestätigen und zu erweitern.
Hinweis E&B: Der folgende farblich markierte Text enthält die Informationen über das wissenschaftliche Vorgehen des Forscherteams. Sie sind wichtig, um die Validität der Untersuchung zu beurteilen, allerdings werden sie den Leserinnen und Lesern, die die hierfür erforderliche Kompetenz nicht besitzen, nicht nützlich sein. Daher wird für primär inhaltlich Interessierte empfohlen, nach der Markierung weiterzulesen und den zusammenfassenden Bericht von E&B zu nutzen, der auch einen Hinweis zum wissenschaftlichen Anspruch der Arbeit enthält.
2 METHODEN
2.1 Teilnehmer
Zu den Teilnehmern gehörten 18 intellektuell hochbegabte Kinder (bestätigt durch neurokognitive Tests), die alle ein ausgeglichenes intellektuelles Profil (mathematisch, verbal, visuomotorisch, Gedächtnis/Konzentration und Urteilsvermögen/Reasoning) aufwiesen, sowie 12 normal entwickelte Kinder. Alle Verfahren standen im Einklang mit der Deklaration von Helsinki und wurden vom Institutional Review Board der Universität von Kalifornien, Los Angeles (UCLA), vor der Aufnahme der Teilnehmer geprüft und genehmigt. Die Teilnehmenden gaben eine schriftliche Einverständniserklärung ab. Sie wurden aus lokalen Schulen und außerschulischen Programmen in Los Angeles rekrutiert, darunter auch Schulen und Programme für besonders begabte Schüler. Die Eignung für diese Studie wurde beim ersten Besuch nach der Durchführung eines standardisierten IQ-Tests (Stanford Binet 5 (Huang-Pollock et al., 2011)) festgestellt. Außergewöhnliche Begabung wurde als IQ ≥ 145 definiert. Außerdem wurden nach Alter, Ethnie und Geschlecht gematchte, gesunde Kinder mit einem IQ ≥90 und≤ 145 rekrutiert, die keine Aufmerksamkeits-, Sprach- oder Lernstörungen aufwiesen. Potenzielle Studienteilnehmer wurden von der Studie ausgeschlossen, wenn eines der folgenden Ausschlusskriterien erfüllt war: Kopfverletzung in der Vorgeschichte, Anfallsleiden oder andere neurologische oder psychiatrische Störungen; frühere oder aktuelle Sprachstörungen, Aufmerksamkeitsdefizit-und Hyperaktivitätsstörungen, Verhaltensstörungen, , Autismus-Spektrum-Störungen oder Drogenabhängigkeit; aktuelle/frühere Einstufung in eine Sonderschulklasse oder ein IQ ≤ 84; Kontraindikationen für MRT (z. B, Metallimplantate, Herzschrittmacher, Zahnspangen oder andere am Kopf befestigte Metalle und Schwangerschaft).
2.2 Demografischer Gruppenvergleich
Demografische Faktoren (z. B. Alter, Bildung) zwischen IG- und TD-Gruppen wurden mittels einseitiger Varianzanalyse (ANOVA) verglichen. Gruppenunterschiede bei dichotomen Faktoren (z. B. Geschlecht, ethnische Zugehörigkeit) wurden mithilfe von Chi-Quadrat-Analysen bewertet. Als Grenzwert für die statistische Signifikanz für diese demografischen Analysen wurde p< .05 verwendet.
2.3 Neuropsychologische Tests
Alle Teilnehmer absolvierten eine standardisierte neuropsychologische Messung (Stanford-Binet 5; Huang-Pollock et al., 2011), die den IQ mit einer hohen Obergrenze testet (IQ= 170) und die Quantifizierung des IQ bei außergewöhnlich intellektuell begabten Teilnehmern ermöglicht.
2.4 MRI-Erfassung
Alle Neuroimaging-Daten wurden mit einer 12-Kanal-Kopfspule an einem 3T Siemens Tim Trio (Siemens Medical Solution, Erlangen, Deutschland) MRT-Scanner im UCLA Staglin IMHRO Center for Cognitive Neuroscience erfasst. Strukturelle MP-RAGE T1-gewichtete Scans wurden mit 120-1,0 mm sagittalen Schichten, FOV = 256 mm (A-P) × 192 mm (FH), Matrix= 256-192, TR= 450,0 ms, TE= 10,0 ms, Flipwinkel= 8, Voxelgröße = 1,0 mm × 0,94 mm × 0,94 mm aufgenommen. Die DTI-Daten wurden unter Verwendung der planaren Einzelschuss-Spin-Echo-Bildgebung (EPI) und unter Verwendung der folgenden Parameter erfasst: TR= 8400 ms; TE= 91 ms; 1282 Matrix, FOV= 256 mm, b= 1000 s/mm2, NEX= 1, 64 Schichten, 2 mm Schichtdicke, 0 mm Skip, PAT= 2. Die Diffusion wurde in 64 Richtungen erfasst (b= 1000 s/mm2) mit 4 Bildern mit b= 0 s/mm2. Alle Bilder wurden vor der Vorverarbeitung und Analyse qualitätskontrolliert und visuell überprüft
2.5 Subkortikale Formverarbeitung und -analyse
Für alle Teilnehmer wurden morphometrische T1-gewichtete anatomische 3D-Scans erstellt. Unter Verwendung der FMRIB Software Library Version 6.0 (Kalbfleisch, 2004) (FSL) wurden die T1-Daten durch ein automatisiertes, modellbasiertes subkortikales Segmentierungsprotokoll (FSL FIRST (Kuhn et al., 2017)) mit einer Randkorrekturmethode geleitet. Die Vertex- und bvar-Dateien der automatisch segmentierten subkortikalen Regionen von Interesse (ROI; die Amygdala, Nucleus accumbens, Caudat, Hippocampus, Pallidum, Putamen und Thalamus) in der linken und rechten Hemisphäre wurden dann visuell auf Qualität geprüft. Die endgültigen Vertex-Dateien für die segmentierten subkortikalen Strukturen wurden sowohl gruppenübergreifend als auch gruppenintern verkettet. Alle automatisch segmentierten ROIs für jeden Teilnehmer wurden am gleichen Standardraum ausgerichtet. Jeder Scheitelpunkt liefert Daten über die Lage der Oberfläche der ROI an demselben Punkt im Raum für jeden Teilnehmer. Zur Durchführung von Gruppenanalysen wurden die Daten jedes Teilnehmers in einem Standardraum registriert, so dass jeder Scheitelpunkt für Gruppenvergleiche auf denselben Punkt im Raum ausgerichtet war. Die mittlere Fläche der Stichprobe wurde als Ziel für diese Ausrichtung verwendet.
Diese skalaren Scheitelwerte wurden dann mit parametrischen statistischen Analysen analysiert, um die Beziehung zwischen der Gruppe (IQ vs. TD) und der Form jeder subkortikalen Struktur zu untersuchen. Diese Analysen wurden für jeden Scheitelpunkt mit Hilfe einer Monte-Carlo-Simulation im FSL-Skript ran-domize (Kyllonen & Christal, 1990, Mills & Tissot, 1995) durchgeführt. Die Regressionen bewerteten die Beziehung zwischen dem radialen Abstand an jedem Scheitelpunkt und den interessierenden Variablen (d.h. Gruppenbezeichnung, IQ). Eine mehrstufige, qualitätskontrollierte Verarbeitungspipeline wurde verwendet, um mehrere Artefaktquellen zu korrigieren, und alle Ergebnisse wurden für Mehrfachvergleiche korrigiert (> .05) unter Verwendung der Falschentdeckungsrate (FDR) (Na et al., 2007).
2.6 Verarbeitung und Analyse der Mikrostruktur der weißen Substanz
Eine routinemäßige DWI-Verarbeitungspipeline wurde mit FSL v 6.0 (Kalbfleisch, 2004) durchgeführt und umfasste die Extraktion des Gehirns, die Korrektur von Wirbelstromverzerrungen sowie die Bewegungs- und Tensoranpassung. DWI-Metriken der Mikrostruktur der weißen Substanz, die fraktionelle Anisotropie (FA) und die mittlere Diffusivität (MD) wurden für jeden Teilnehmer berechnet. Die Analysen wurden mit Hilfe der Tract Based Spatial Statistics (TBSS) (Navas-Sánchez et al., 2014) durchgeführt, einer Methode, die eine voxelweise statistische Abfrage der DWI-Metriken entlang der WM-Trakte von Interesse (TOI) ermöglicht. Die TOI wurden aus dem Atlas der weißen Materie der John Hopkins University (Neihart et al., 2002) abgeleitet und umfassten bilateral: die anteriore thalamische Strahlung (ATR), die inferioren fronto- okzipitalen Faszikel (IFO), den inferioren longitudinalen Faszikulus (ILF), den superioren longitudinalen Faszikulus (SLF), den Fasciculus uncinatus (UNC), den cingulären Bun- den (CGC), die weiße Substanz des Hippocampus (HC) sowie die Zangen major und die Zangen minor. Parametrische statistische Analysen untersuchten die Unterschiede zwischen den Gruppen in der WM-Mikrostruktur (FA und MD). Korrelationsanalysen untersuchten die Beziehung zwischen IQ und FA/MD. Die TBSS-Ergebnisse wurden für Mehrfachvergleiche mit threshold free cluster enhancement-TFCE (O’Boyle, 2008) korrigiert.
2.7 Volumetrische Bestätigung der expliziten/impliziten Heuristik anhand eines externen Datensatzes
Die aktuelle Stichprobe enthielt keine Kinder mit unterdurchschnittlichen bis beeinträchtigten IQ-Werten oder atypischen neurologischen Entwicklungsbedingungen wie Aufmerksamkeitsdefizit-Hyperaktivitätsstörung (ADHS) oder Autismus. Daher wollten wir die explizite/implizite Rationalisierungsheuristik in einem externen, großen Datensatz testen, der dieses breitere Spektrum an Entwicklungsverläufen umfasst. Daher wurde das Adolescent Brain and Cognitive Development (ABCD)-Projekt (UCLA PI: Bookheimer) als externer Datensatz verwendet, um zu versuchen, die aus der Analyse der begabten Jugendlichen abgeleitete Heuristik der expliziten und impliziten Rationalität zu validieren (beschrieben in Abschnitt 3). Alter, Geschlecht, globaler kognitiver Funktionswert aus der Toolbox des National Institute of Health (NIH) sowie das Volumen der subkortikalen Strukturen wurden von allen verfügbaren ABCD-Teilnehmern zusammengestellt. Das Volumen der (expliziten Gedächtnis-)Hirnregionen, die in der IG-Gruppe signifikant größer waren, wurde summiert, um die explizite Gedächtnis-Hirnkomponente zu bilden. Die Volumina der (impliziten Gedächtnis-)Hirnregionen, die in der TD-Gruppe größer waren, wurden summiert, um die implizite Gedächtnis-Hirnkomponente zu bilden. Dann wurde die explizite/implizite Verhältnisvariable berechnet, indem die explizite Gedächtnis-Hirnkomponente durch die implizite Gedächtnis-Hirnkomponente geteilt wurde. Anschließend wurde eine Korrelationsanalyse berechnet, um die Beziehung zwischen diesem expliziten/impliziten Gedächtnis-Gehirn-Verhältnis und der globalen kognitiven Leistung, die mit der NIH-Toolbox gemessen wurde, zu bewerten. In Anbetracht des Formats, in dem die ABCD-Daten zur Verfügung gestellt werden (d. h. extrahierte Werte in einer Tabellenkalkulation und keine MRT-Rohdaten), war es uns nicht möglich, eine Formanalyse durchzuführen. Wir waren jedoch in der Lage, Volumenanalysen durchzuführen. Wichtig ist, dass Formanalysen regionalspezifischer und empfindlicher sind als Volumenanalysen (Rogers, 1986). Effekte, die in der Volumenanalyse gefunden werden, sind auch in der Formanalyse vorhanden. Die Ergebnisse der Formanalyse können jedoch vorhanden sein, wenn die Volumenanalyse nicht empfindlich genug ist, um Veränderungen zu erkennen (Rota et al., 2009). Daher sollte eine erfolgreiche Replikation der Heuristik unter Verwendung der volumetrischen ABCD-Daten Ergebnisse der Formanalyse aus der ursprünglichen Stichprobe hier effektiv wiederholen.
3 ERGEBNISSE
3.1 Vergleich der demografischen Gruppen
Die IG- und die TD-Gruppe unterschieden sich nicht signifikant in Bezug auf Alter, Schulbildung, ethnische Zugehörigkeit oder Geschlecht (alle p> .05). Die IG-Gruppe (153± 11,4) hatte einen signifikant höheren IQ als die TD-Gruppe (128± 10,9, p< .01; Tabelle 1). Die IG-Gruppe setzte sich aus „ausgewogenen“ intellektuellen Profilen zusammen. Alle begabten Teilnehmer befanden sich auf der Stufe Superior Adult 1 oder höher für mathematische und sprachliche Fähigkeiten, mit einer Ausnahme. Ein begabter Teilnehmer lag bei „Wortschatz und Redegewandtheit“ auf der Stufe „Überdurchschnittlich“ und bei „Arithmetisches Denken“ auf der Stufe „Überdurchschnittlich Erwachsener 1“. Derselbe Teilnehmer erreichte auch in den Bereichen Urteilsvermögen und logisches Denken die Stufe Superior Adult 3. Daher schienen alle IG-Teilnehmer in ihren mathematischen und sprachlichen Fähigkeiten ausgeglichen zu sein.
3.2 Subkortikale Form
Signifikant mehr Scheitelpunkte wurden in den bilateralen Hippocampi und dem rechten Putamen von IG gefunden, während bei 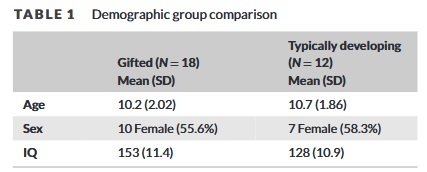 TD größere Scheitelpunkte in der linken Amygdala, dem rechten Caudate und dem bilateralen Nucleus accumbens gefunden wurden. Speziell IG wies größere Scheitelpunkte im Kopf des linken HC, im Körper des rechten HC und im bilateralen HC-Schwanz sowie in den hinteren Regionen des rechten Putamen auf. TD wies größere Scheitelpunkte im gesamten bilateralen Accumbens auf, hauptsächlich im Körper des rechten Caudatus und in den zentromedianen Kernen der linken Amygdala. In der gesamten Stichprobe korrelierte der IQ mit größeren Scheitelpunkten im bilateralen HC-Schwanz (Abbildung 2).
TD größere Scheitelpunkte in der linken Amygdala, dem rechten Caudate und dem bilateralen Nucleus accumbens gefunden wurden. Speziell IG wies größere Scheitelpunkte im Kopf des linken HC, im Körper des rechten HC und im bilateralen HC-Schwanz sowie in den hinteren Regionen des rechten Putamen auf. TD wies größere Scheitelpunkte im gesamten bilateralen Accumbens auf, hauptsächlich im Körper des rechten Caudatus und in den zentromedianen Kernen der linken Amygdala. In der gesamten Stichprobe korrelierte der IQ mit größeren Scheitelpunkten im bilateralen HC-Schwanz (Abbildung 2).
3.3 Mikrostruktur der weißen Substanz
Die FA in der weißen Substanz der rechten ATR und des rechten HC-Schwanzes (d. h. Fornix) war in IG-Gruppe im Vergleich zu den Kontrollen signifikant höher (.001< p < .05). Die MD war in der IG-Gruppe im Vergleich zu den Kontrollen in der bilateralen ATR, HC und UNC signifikant niedriger. Insbesondere wurden MD-Ergebnisse in einem höheren Prozentsatz der rechten im Vergleich zur linken Hemisphäre gefunden. Die MD-Ergebnisse wurden auch in verschiedenen Regionen je nach Hemisphäre gefunden. Bei den HC fanden sich die Ergebnisse in der rechten Hemisphäre in der Nähe des HC-Schwanzes (d. h. Fornix) und in der linken Hemisphäre im HC-Körper (d. h. in den CA-Regionen). Bei den UNC waren die Ergebnisse in der linken Hemisphäre geringer. In der gesamten Stichprobe korrelierte die FA signifikant positiv mit dem IQ in der bilateralen (rechte Hemisphäre mehr als linke) ATR und der rechten HC. Darüber hinaus korrelierte die MD negativ mit dem IQ in der bilateralen (rechts stärker als links) ATR, der linken HC, der linken UNC, dem linken cingulären Bündel und der linken ILF (Abbildungen 1 und 2).
3.4 Bestätigung der expliziten/impliziten Heuristik anhand eines externen Datensatzes
Der ABCD-Datensatz umfasste 7652 Teilnehmer (Durchschnittsalter= 119 ± 7,5 Monate; 62,9 % weiblich) mit allen für diese Studie erforderlichen Daten. Das Verhältnis explizites/implizites Gedächtnis war signifikant positiv mit der globalen kognitiven Leistung (r = 0,23, p < 0,05) sowie der quadratischen globalen kognitiven Leistung (r = .022, p < .05) verbunden (Abbildung 4). Eine schrittweise hierarchische Regression mit linearer und quadratischer globaler kognitiver Leistung ergab ein endgültiges Modell [F (2, 7652)= 4.50, = .011) einschließlich der quadratischen globalen kognitiven Leistung (β= 0,24, p = .04) als signifikanter Prädiktor für das explizite/implizite Verhältnis. Da die Formanalyse empfindlicher ist als die Volumenanalyse, sollte dieses Ergebnis bei Verwendung der Volumetrie im ABCD-Datensatz auch auf die Formanalyse übertragbar sein (was aufgrund des Formats der ABCD-Daten nicht möglich war).
Eine ähnliche Regression wurde unter Verwendung der Messungen der globalen Kognition durchgeführt, um eine zusammengesetzte Variable vorherzusagen, die aus der FA der weißen Substanz der ATR und der HC bestand. Es gab eine signifikante quadratische Beziehung [F (2, 7562)= 13.17, p< .0001] zwischen der globalen Kognition (β= -0.034, p < .001) und die zusammengesetzte Variable, die aus der FA der rechten ATR und der weißen Substanz der HC bestand. Schließlich wurde durch eine schrittweise Regression eine zusammengesetzte Variable vorhergesagt, die sich aus der MD der UNC, der weißen Substanz der HC und der ATR zusammensetzte. Der quadratische globale kognitive Score (β= 0,12, p< .001) sagte diese zusammengesetzte MD-Variable signifikant voraus [F (2, 7562)= 15,31, p< .0001].
4 DISKUSSION
Diese Untersuchung ergab, dass es eine doppelte Dissoziation zwischen der Größe und der Konnektivität von zwei separaten Gedächtnissystemen gibt, wenn man intellektuell begabte Kinder und ihre TD-Gegenstücke vergleicht. Im Vergleich zur TD-Gruppe (typically developing) wiesen die begabten Kinder größere subkortikale Strukturen und eine stärker vernetzte mikrostrukturelle Organisation der weißen Substanz zwischen diesen Strukturen in Regionen auf, die mit explizitem Gedächtnis und IQ in Verbindung stehen: bilateraler Hippocampus und rechtes Putamen. Insbesondere das Putamen, wahrscheinlich über integrierende Verbindungen mit dem präfrontalen Kortex, und Unterregionen des Hippocampus, die mit neuem Lernen und Informationsintegration (Gyrus dentatus und CA3) in Verbindung stehen, waren bei intellektuell begabten Kindern größer (Abbildungen 1 und 2). Die ausgeprägte Integrität der Verbindungen der weißen Substanz zwischen diesen Regionen entspricht einer anatomischen Grundlage für das angeborene Denkverhalten intellektuell begabter Kinder, explizite Informationen schnell und effizient zu lernen, zu integrieren und zu nutzen. Darüber hinaus könnten diese Ergebnisse mit der Neigung begabter Kinder zusammenhängen, ein höheres Maß an intrinsischer Motivation zu zeigen, zu lesen, zu denken und Zeit allein zu verbringen. Interessanterweise wiesen typisch entwickelte Kinder mehr verbundene, größere subkortikale Strukturen in Regionen auf, die mit dem impliziten Gedächtnis in Verbindung gebracht werden: Striatum (d. h. Caudat, Nucleus accumbens) und Amygdala (Abbildung 2; Haier et al., 1988).
Diese doppelte Dissoziation deutet darauf hin, dass intellektuell begabte und sich normal entwickelnde Kinder möglicherweise von Natur aus unterschiedliche neurologische Entwicklungsverläufe haben, die durch unterschiedliche Lernstrategien erkennbar werden.
Diese Erkenntnis hat uns dazu veranlasst, uns ein breites Spektrum von Intelligenztypen vorzustellen, die u.a. unterschiedliche Verhältnisse zwischen impliziter und expliziter Systementwicklung aufweisen. Das explizite und das implizite Gedächtnis belegen unterschiedliche Regionen des Gehirns und bilden unabhängige, aber miteinander verbundene Lernnetze. Aufgrund der begrenzten Ressourcen und des begrenzten Platzes im Gehirn kann es zu einem Kompromiss zwischen der Entwicklung expliziter und impliziter Gedächtnissysteme kommen. Die Unterschiede, die wir gefunden haben, spiegeln möglicherweise eine breite Streuung der Gedächtnissystemvariation zwischen TD und IG wider, wobei TD und IG die Pole / die funktionellsten Nischen darstellen (Abbildung 3).
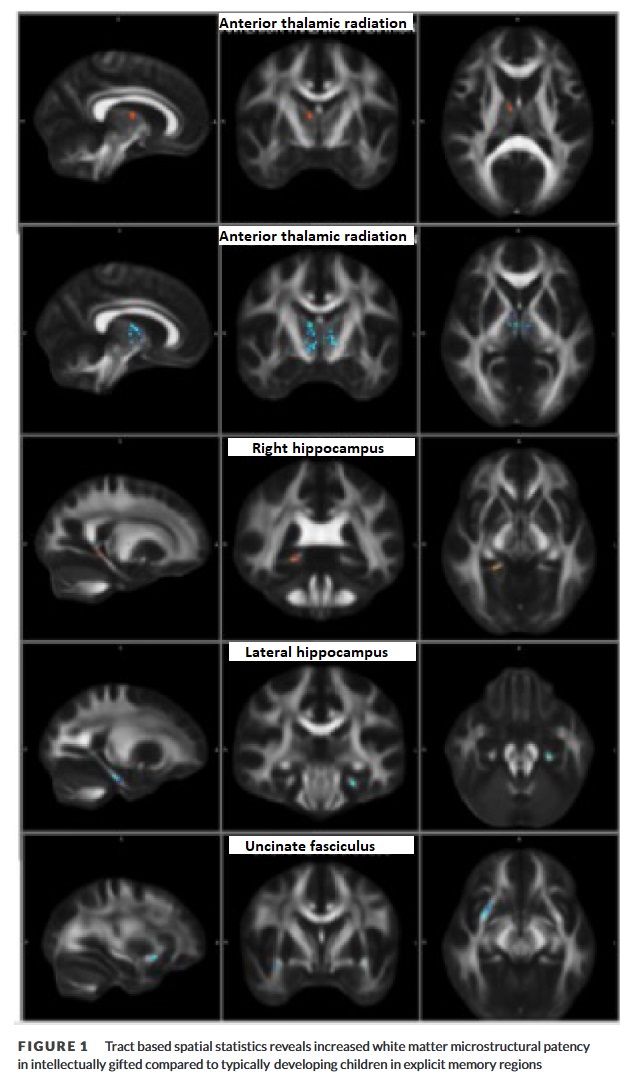
Die Befunde aus dem Gehirn eines mathematischen Wunderkinds wurden zuvor als Folge der hocheffiziente Kodierung und Abfrage episodischer Erinnerungen (im expliziten Gedächtnis, Eck) interpretiert (Scharnowski et al., 2015). Damit dies zutrifft, müssten sowohl Strukturen des episodischen Gedächtnisses als auch Systeme des Frontallappens, die an der Aufmerksamkeit und dem Abruf beteiligt sind, vorhanden sein. Die vorliegende Studie zeigt genau das: große und robuste Verbindungen zwischen den Strukturen des episodischen Gedächtnisses und den Bahnen der weißen Substanz im Frontallappen (z. B. ATR). – Der Befund der ATR steht auch im Einklang mit einer Vielzahl früherer Arbeiten, in denen festgestellt wurde, dass hohe Intelligenz mit der Aktivierung präfrontaler und zingulärer Hirnnetzwerke verbunden ist (Kuhn et al., 2017, Schmithorst et al., 2005) sowie früheren Arbeiten, in denen festgestellt wurde, dass mathematisch begabte Gehirne eine höhere FA im Corpus callosum und in den Assoziationsbahnen aufweisen, die den Frontallappen mit den Basalganglienstrukturen, einschließlich des ATR, und den 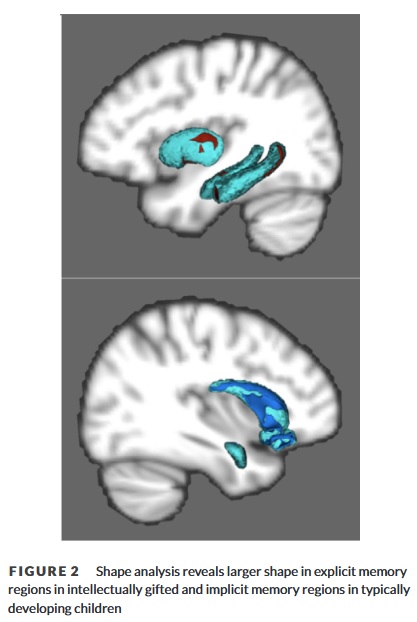 fronto-temporalen/parietalen Bahnen, einschließlich des UNC, verbinden (Shaw et al., 2006). Die hier gefundenen verstärkten bilateralen Hirnregionen stehen im Einklang mit früheren Erkenntnissen, die darauf hindeuten, dass einzigartige bilaterale Hirnregionen von mathematisch frühreifen Kindern während mentaler Rotationsaufgaben aktiviert werden (Squire et al., 1993, Weiskopf, 2012). Darüber hinaus steht die Tendenz zur Lateralisierung der Ergebnisse im Einklang mit früheren Erkenntnissen, die darauf hindeuten, dass die Lateralität der Hemisphären, insbesondere der rechten Hemisphäre, sowie eine verbesserte Koordination innerhalb und zwischen den Hirnregionen eine wichtige neuronale Grundlage für intellektuelle Begabung darstellen (O’Boyle et al., 1995). Schließlich stehen die Ergebnisse der gesamten Stichprobe, die darauf hindeuten, dass der IO mit den DTI-Metriken in der ATR, HC und UNC korreliert ist, im Einklang mit früheren Ergebnissen einer Studie über mathematische Begabung (Shaw et al., 2006), in der Korporalkallosum, Fornix und DTI-Metrik des Assoziationstrakts mit der Intelligenz korreliert wurden. Diese Beziehung zwischen weißer Substanz und IO wird möglicherweise zumindest teilweise von der Genfamilie Plexin bestimmt, die kürzlich in einer genomweiten Assoziationsstudie zur Vorhersage des IO herangezogen wurde (Winberg et al., 1998). Es ist bekannt, dass Plexine mit der Führung von sich entwickelnden Axonen (Worzfeld & Offermanns, 2014), der neuronalen Konnektivität (Zabaneh et al., 2018) und der Regeneration (Zhang et al., 2017) in Verbindung stehen und somit mit den Ergebnissen der weißen Substanz in unserer IG-Gruppe in Zusammenhang stehen könnten.
fronto-temporalen/parietalen Bahnen, einschließlich des UNC, verbinden (Shaw et al., 2006). Die hier gefundenen verstärkten bilateralen Hirnregionen stehen im Einklang mit früheren Erkenntnissen, die darauf hindeuten, dass einzigartige bilaterale Hirnregionen von mathematisch frühreifen Kindern während mentaler Rotationsaufgaben aktiviert werden (Squire et al., 1993, Weiskopf, 2012). Darüber hinaus steht die Tendenz zur Lateralisierung der Ergebnisse im Einklang mit früheren Erkenntnissen, die darauf hindeuten, dass die Lateralität der Hemisphären, insbesondere der rechten Hemisphäre, sowie eine verbesserte Koordination innerhalb und zwischen den Hirnregionen eine wichtige neuronale Grundlage für intellektuelle Begabung darstellen (O’Boyle et al., 1995). Schließlich stehen die Ergebnisse der gesamten Stichprobe, die darauf hindeuten, dass der IO mit den DTI-Metriken in der ATR, HC und UNC korreliert ist, im Einklang mit früheren Ergebnissen einer Studie über mathematische Begabung (Shaw et al., 2006), in der Korporalkallosum, Fornix und DTI-Metrik des Assoziationstrakts mit der Intelligenz korreliert wurden. Diese Beziehung zwischen weißer Substanz und IO wird möglicherweise zumindest teilweise von der Genfamilie Plexin bestimmt, die kürzlich in einer genomweiten Assoziationsstudie zur Vorhersage des IO herangezogen wurde (Winberg et al., 1998). Es ist bekannt, dass Plexine mit der Führung von sich entwickelnden Axonen (Worzfeld & Offermanns, 2014), der neuronalen Konnektivität (Zabaneh et al., 2018) und der Regeneration (Zhang et al., 2017) in Verbindung stehen und somit mit den Ergebnissen der weißen Substanz in unserer IG-Gruppe in Zusammenhang stehen könnten.
Diese erste Stichprobe umfasste keine Kinder mit unterdurchschnittlichen oder beeinträchtigten IO-Werten oder atypischen neurologischen Entwicklungsbedingungen wie ADHS oder Autismus. Diese Gruppen können am anderen Ende des Spektrums liegen, wo das explizite oder implizite Gedächtnis unter- oder überentwickelt sein kann, was sich nachteilig auswirken kann. Daher wiederholten wir diese Analyse in einer viel größeren Stichprobe, die auch Kinder mit unterdurchschnittlichen bis beeinträchtigten IO-Werten und einigen Entwicklungsunterschieden (z. B. ADHS) umfasste. Wir testeten diese explizite/implizite Heuristik anhand eines großen, externen Datensatzes: dem ABCD-Projekt (N= 7652). In mit unserer Hypothese fanden wir eine signifikante quadratische Beziehung zwischen der expliziten/impliziten Heuristik und dem IO. Diese Replikation unserer ursprünglichen Ergebnisse deutet darauf hin, dass es in der Tat ein entwicklungsbedingtes Gleichgewicht zwischen impliziten und expliziten Gedächtnissystemen geben kann, das sich phänotypisch in unterschiedlichen kognitiven Profilen ausdrücken kann. Zukünftige Forschungen könnten die relative Entwicklung von impliziten und expliziten Strukturen in anderen Populationen untersuchen, wie dies bei neurodegenerativen Störungen wie der Alzheimer-Krankheit (degenerierende explizite Gedächtnisanatomie und – fähigkeiten bei erhaltenem impliziten Gedächtnis) und der Parkinson-Krankheit (degenerierende implizite Gedächtnisanatomie und -fähigkeiten bei erhaltenem expliziten Gedächtnis) eingehend untersucht wurde.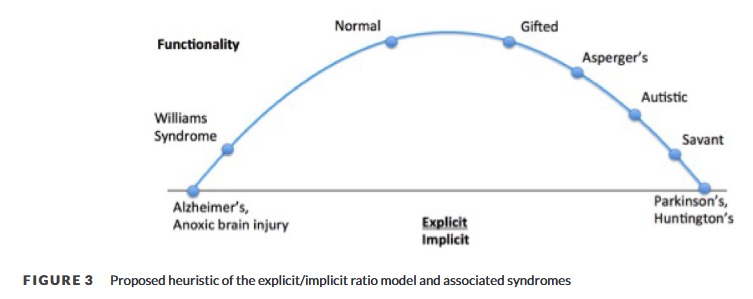
Die Identifizierung der strukturellen und funktionalen Merkmale von Hochbegabung wird uns helfen, die tieferen Systeme zu verstehen, die es Kindern ermöglichen, innerhalb und außerhalb der Schule lernen. Sowohl implizites als auch explizites Lernen sind wichtig, und Defizite in beiden Bereichen können zu sozialen, akademischen und beruflichen Schwierigkeiten führen. Obwohl „begabte“ Kinder in IO-Tests gut abschneiden, leiden viele von ihnen auch an Lernstörungen, die sie bei nicht expliziten Lernaufgaben behindern, und die meisten TD-Kinder könnten von einer weiteren Entwicklung des expliziten Lernens profitieren (Hayden et al., 2020). Diese Studie gibt Aufschluss darüber, wie künftige Interventionen explizite oder implizite Systeme fördern können, um das Lernen im gesamten Entwicklungsspektrum zu maximieren. Möglicherweise wird die Bildgebung des Gehirns es uns in Zukunft ermöglichen, pädagogische Interventionen auf der Grundlage der individuellen Gehirnstruktur zu entwickeln, um eine gesunde intellektuelle und verhaltensbezogene Entwicklung zu fördern. Empirisch begründete kognitive Trainingsverfahren,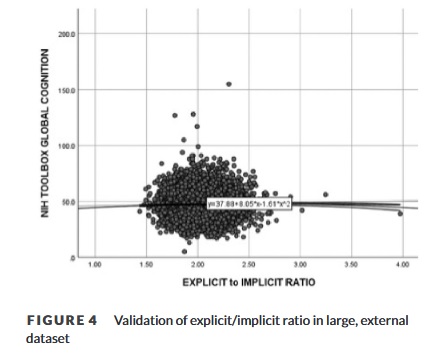 wie das Program for Education and Enrishment of Relational Skills (PEERS; Laugeson et al., 2012), könnten bei der Förderung von TD-Kindern, begabten Kindern und gefährdeten klinischen Populationen (z. B. Autismus-Spektrum-Störungen, Lernstörungen, Aufmerksamkeitsdefizit-Hyperaktivitätsstörungen und intellektuelle Behinderungen) helfen und es Bildungseinrichtungen ermöglichen, Kinder aller Lerntypen und Fähigkeiten zu unterstützen. Wir hoffen, dass diese Studie zu weiteren Untersuchungen über die Strukturen anregt, die implizites und explizites Lernen erleichtern, über ihre Rolle bei Entwicklungsunterschieden und über mögliche zukünftige Lerntherapien.
wie das Program for Education and Enrishment of Relational Skills (PEERS; Laugeson et al., 2012), könnten bei der Förderung von TD-Kindern, begabten Kindern und gefährdeten klinischen Populationen (z. B. Autismus-Spektrum-Störungen, Lernstörungen, Aufmerksamkeitsdefizit-Hyperaktivitätsstörungen und intellektuelle Behinderungen) helfen und es Bildungseinrichtungen ermöglichen, Kinder aller Lerntypen und Fähigkeiten zu unterstützen. Wir hoffen, dass diese Studie zu weiteren Untersuchungen über die Strukturen anregt, die implizites und explizites Lernen erleichtern, über ihre Rolle bei Entwicklungsunterschieden und über mögliche zukünftige Lerntherapien.
Teil 2 im Original
CONFLICT OF INTEREST The authors report no conflicts of interest.
PEER REVIEW The peer review history for this article is available at https://publons.com/publon/10.1002/brb3.2348
ACKNOWLEDGMENTS This work was supported by the University of California, Los Angeles Staglin Center for Cognitive Neuroscience (PI: Susan Y. Bookheimer, PhD), the Tennenbaum Center for the Biology of Creativity and Joaquin Fuster Foundation. The authors thank Monika Polcynska‐Bledsos for her contribution to the manuscript preparation process.
Kuhn, T. , Blades, R. , Gottlieb, L. , Knudsen, K. , Ashdown, C. , Martin‐Harris, L. , Ghahremani, D. , Dang, B. H. , Bilder, R. M. , & Bookheimer, S. Y. (2021). Neuroanatomical differences in the memory systems of intellectual giftedness and typical development. Brain and Behavior, 11, e2348. 10.1002/brb3.2348
DATA AVAILABILITY STATEMENT The data that support the findings of this study are available from the corresponding author upon reasonable request.
Associated Data This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement The data that support the findings of this study are available from the corresponding author upon reasonable request.
Articles from Brain and Behavior are provided here courtesy of Wiley
REFERENCES
- Ali, N. , Na’aya, H. U. , & Ngadda, H. (2003). Cancer of the gall bladder: A report of three cases and a review of the literature. Nigerian Journal of Medicine, 12, 232–235. [PubMed] [Google Scholar]
- Athanasakis, E. , Licastro, D. , Faletra, F. , Fabretto, A. , Dipresa, S. , Vozzi, D. , Morgan, A. , d’Adamo, A. P. , Pecile, V. , Biarnés, X. , & Gasparini, P. , (2014). Next generation sequencing in nonsyndromic intellectual disability: From a negative molecular karyotype to a possible causative mutation detection. American Journal of Medical Genetics A, 164, 170–176. [DOI] [PubMed] [Google Scholar]
- Colom, R. , Rebollo, I. , Palacios, A. , Juan‐Espinosa, M. , & Kyllonen, P. C. (2004). Working memory is (almost) perfectly predicted by g. Intelligence, 32, 277–296. [Google Scholar]
- Davidson, J. E. (1986). The role of insight in giftedness. Conceptions of Giftedness, 201–222. [Google Scholar]
- Desco, M. , Navas‐Sanchez, F. J. , Sanchez‐González, J. , Reig, S. , Robles, O. , Franco, C. , Guzmán‐De‐Villoria, J. A. , García‐Barreno, P. , & Arango, C. (2011). Mathematically gifted adolescents use more extensive and more bilateral areas of the fronto‐parietal network than controls during executive functioning and fluid reasoning tasks. Neuroimage, 57(1), 281–292. [DOI] [PubMed] [Google Scholar]
- Duncan, J. , Seitz, R. J. , Kolodny, J. , Bor, D. , Herzog, H. , Ahmed, A. , Newell, F. , & Emslie, H. (2000). A neural basis for general intelligence. Science, 289, 457–460. [DOI] [PubMed] [Google Scholar]
- Geake, J. , & Hansen, P. (2005). Neural correlates of intelligence as revealed by fMRI of fluid analogies. Neuroimage, 26, 555–564. [DOI] [PubMed] [Google Scholar]
- Gerig, G. , Styner, M. , Shenton, M. E. , & Lieberman, J. A. (2001). Shape versus size: Improved understanding of the morphology of brain structures. In International conference on medical image computing and computer‐assisted intervention (pp. 24–32). Springer. [Google Scholar]
- Gonring, K. , Gerdes, A. , & Gardner, D. (2017). Program for the education and enrichment of relational skills: Parental outcomes with an ADHD sample. Child & Family Behavior Therapy, 39(1), 19–42. [Google Scholar]
- Gray, J. R. , Chabris, C. F. , & Braver, T. S. (2003). Neural mechanisms of general fluid intelligence. Nature Neuroscience, 6, 316–322. [DOI] [PubMed] [Google Scholar]
- Haier, R. J. , Siegel, B. V. Jr. , Nuechterlein, K. H. , Hazlett, E. , Wu, J. C. , Paek, J. , Browning, H. L. , & Buchsbaum, M. S. (1988). Cortical glucose metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography. Intelligence, 12, 199–217. [Google Scholar]
- Hayden, P. J. , Iacobelli, S. , Perez‐Simon, J. A. , van Biezen, A. , Minnema, M. , Nittyvuopio, R. , Schonland, S. , Meijer, E. , Blaise, D. , Milpied, N. , Marquez‐Malaver, F. J. , Veelken, J. H. , Maertens, J. , Michallet, M. , Cammenga, J. , N’Guyen, S. , Niederwieser, D. , Hunault‐Berger, M. , Bourhis, J. H. , … Kroger, N. (2020). Conditioning‐based outcomes after allogeneic transplantation for myeloma following a prior autologous transplant (1991–2012) on behalf of EBMT CMWP. European Journal of Haematology, 104, 181–189. [DOI] [PubMed] [Google Scholar]
- Huang‐Pollock, C. L. , Maddox, W. T. , & Karalunas, S. L. (2011). Development of implicit and explicit category learning. Journal of Experimental Child Psychology, 109, 321–335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalbfleisch, M. L. (2004). Functional neural anatomy of talent. The Anatomical Record Part B, 277, 21–36. [DOI] [PubMed] [Google Scholar]
- Kuhn, T. , Schonfeld, D. , Sayegh, P. , Arentoft, A. , Jones, J. D. , Hinkin, C. H. , Bookheimer, S. Y. , & Thames, A. D. (2017). The effects of HIV and aging on subcortical shape alterations: A 3D morphometric study. Human Brain Mapping, 38(2), 1025–1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyllonen, P. C. , & Christal, R. E. (1990). Reasoning ability is (little more than) working‐memory capacity? Intelligence, 14, 389–433. [Google Scholar]
- Laugeson, E. A. , Frankel, F. , Gantman, A. , Dillon, A. R. , & Mogil, C. (2012). Evidence‐based social skills training for adolescents with autism spectrum disorders: The UCLA PEERS program. Journal of autism and developmental disorders, 42(6), 1025–1036. [DOI] [PubMed] [Google Scholar]
- Mills, C. J. , & Tissot, S. L. (1995). Identifying academic potential in students from under‐represented populations: Is using the Ravens Progressive Matrices a good idea? Gifted Child Quarterly, 39, 209–217. [Google Scholar]
- Na, H. S. , Hong, S. J. , Yoon, H.J. , Maeng, J. H. , Ko, B. M. , Jung, I. S. , Ryu, C. B. , Kim, J. O. , Cho, J. Y. , Lee, J. S. , Lee, M. S. , Shim, C.S. , & Kim, B. S. (2007). Eradication rate of first‐line and second‐line therapy for Helicobacter pylori infection, and reinfection rate after successful eradication. The Korean Journal of Gastroenterology, 50, 170–175. [PubMed] [Google Scholar]
- Navas‐Sánchez, F. J. , Alemán‐Gómez, Y. , Sánchez‐Gonzalez, J. , Guzmán‐De‐Villoria, J. A. , Franco, C. , Robles, O. , Arango, C. , & Desco, M. (2014). White matter microstructure correlates of mathematical giftedness and intelligence quotient. Human Brain Mapping, 35(6), 2619–2631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neihart, M. , Reis, S. M. , Robinson, N. , & Moon, S. (2002). The social and emotional development of gifted children: What do we know? Sourcebooks, Inc. [Google Scholar]
- O’Boyle, M. W. (2008). Mathematically gifted children: Developmental brain characteristics and their prognosis for well‐being. Roeper Review, 30(3), 181–186. [Google Scholar]
- O’Boyle, M. W. , Alexander, J. E. , & Benbow, C. P. (1991). Enhanced right hemisphere activation in the mathematically precocious: A preliminary EEG investigation. Brain and Cognition, 17, 138–153. [DOI] [PubMed] [Google Scholar]
- O’Boyle, M. W. , & Benbow, C. P. (1990). Enhanced right hemisphere involvement during cognitive processing may relate to intellectual precocity. Neuropsychologia, 28, 211–216. [DOI] [PubMed] [Google Scholar]
- O’Boyle, M. W. , Benbow, C. P. , & Alexander, J. E. (1995). Sex differences, hemispheric laterality, and associated brain activity in the intellectually gifted. Developmental Neuropsychology, 11(4), 415–443. [Google Scholar]
- O’Boyle, M. W. , Cunnington, R. , Silk, T. J. , Vaughan, D. , Jackson, G. , Syngeniotis, A. , & Egan, G. F. (2005). Mathematically gifted male adolescents activate a unique brain network during mental rotation. Cognitive Brain Research, 25(2), 583–587. [DOI] [PubMed] [Google Scholar]
- Packard, M. G. , & Knowlton, B. J. (2002).Learning and memory functions of the basal ganglia. Annual Review of Neuroscience, 25, 563–593. [DOI] [PubMed] [Google Scholar]
- Pesenti, M. , Zago, L. , Crivello, F. , Mellet, E. , Samson, D. , & Duroux, B. (2001). Mental calculation in a prodigy is sustained by right prefrontal and medial temporal areas. Nature Neuroscience, 4, 103–107. [DOI] [PubMed] [Google Scholar]
- Rogers, K. B. (1986). Do the gifted think and learn differently?: A review of recent research and its implications for instruction. Citeseer. [Google Scholar]
- Rota, G. , Sitaram, R. , Veit, R. , Erb, M. , Weiskopf, N. , Dogil, G. , & Birbaumer, N. (2009). Self‐regulation of regional cortical activity using real‐time fMRI: The right inferior frontal gyrus and linguistic processing. Human Brain Mapping, 30, 1605–1614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scharnowski, F. , Veit, R. , Zopf, R. , Studer, P. , Bock, S. , Diedrichsen, J. , Goebel, R. , Mathiak, K. , Birbaumer, N. , & Weiskopf, N. (2015). Manipulating motor performance and memory through real‐time fMRI neurofeedback. Biological Psychology, 108, 85–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmithorst, V. J. , Wilke, M. , Dardzinski, B. J. , & Holland, S. K. (2005). Cognitive functions correlate with white matter architecture in a normal pediatric population: A diffusion tensor MRI study. Human Brain Mapping, 26, 139–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw, P. , Greenstein, D. , Lerch, J. , Clasen, L. , Lenroot, R. , Gogtay, N. , Evans, A. , Rapoport, J. , & Giedd, J. (2006). Intellectual ability and cortical development in children and adolescents. Nature, 440, 676–679. [DOI] [PubMed] [Google Scholar]
- Squire, L. R. , Knowlton, B. , & Musen, G. (1993). The structure and organization of memory. Annual Review of Psychology, 44, 453–495. [DOI] [PubMed] [Google Scholar]
- Weiskopf, N. (2012). Real‐time fMRI and its application to neurofeedback. Neuroimage, 62, 682–692. [DOI] [PubMed] [Google Scholar]
- Winberg, M. L. , Noordermeer, J. N. , Tamagnone, L. , Comoglio, P. M. , Spriggs, M. K. , Tessier‐Lavigne, M. , & Goodman, C. S. (1998). Plexin A is a neuronal semaphorin receptor that controls axon guidance. Cell, 95, 903–916. [DOI] [PubMed] [Google Scholar]
- Worzfeld, T. , & Offermanns, S. (2014). Semaphorins and plexins as therapeutic targets. Nature Reviews Drug Discovery, 13, 603–621. [DOI] [PubMed] [Google Scholar]
- Zabaneh, D. , Krapohl, E. , Gaspar, H. A. , Curtis, C. , Lee, S. H. , Patel, H. , Newhouse, S. , Wu, H. M. , Simpson, M. A. , Putallaz, M. , Lubinski, D. , Plomin, R. , & Breen, G. (2018). A genome‐wide association study for extremely high intelligence. Molecular Psychiatry, 23(5), 1226–1232. [DOI] [PMC free article] [PubMed] [Google Scholar]
Zhang, L. , Gan, J. Q. , & Wang, H. (2017). Neurocognitive mechanisms of mathematical giftedness: A literature review. Applied Neuropsychology: Child, 6(1), 79–94. [DOI] [PubMed] [Google Scholar